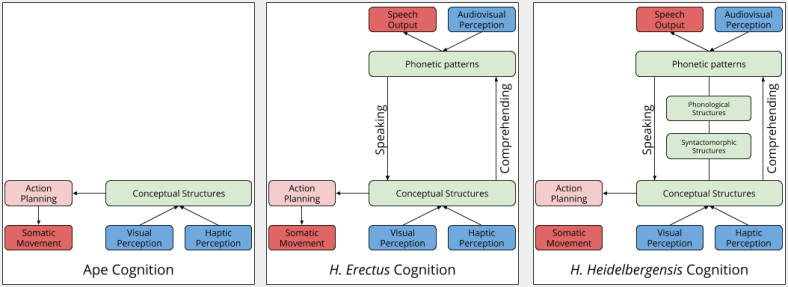
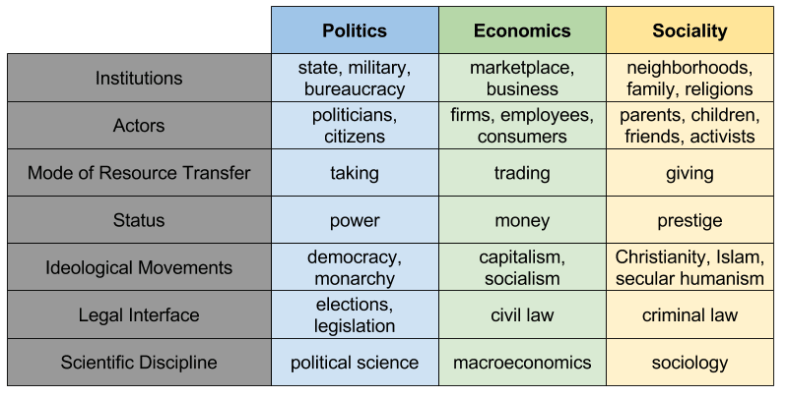
The Three Sphere Hypothesis
Most people agree that human societies operate in different contexts: markets, governments, and communities. The Three Sphere Hypothesis holds that this trichotomy is fundamental and exhaustive of social space. What’s more, these spheres interact. Neither markets nor governments nor communities can be analyzed thoroughly without understanding their dependence upon, and their effects upon, the others.
[Excerpt] Intellectual History of the Hypothesis
Source: Wicks (2009). A Model of Dynamic Balance among the Three Spheres of Society
Social scientists – including economists – as well as journalists and others, often refer to “the economic, political, and social conditions” underlying any particular situation, but usually without any further analysis of what these terms imply, and how they relate to each other.
Apparent references to these three spheres pop up – in both popular and technical literature – almost everywhere. It can be a fun game, like “whack-a-mole”:
- Where and how will the three spheres “pop up” in this or that text?
- And, given any set of three social attributes that do “pop up”, can they be seen in some way as representing the three spheres?
Etzioni (1996:122) speaks of “three different conditions: paid, coerced, or convinced”; Etzioni (1988) explores motivations in the community sphere at length.
Personalist economics, based on Catholic theology, also recognizes three organizing principles: competition, intervention, and cooperation (Jonish and Terry, 1999:465-6; O’Boyle, 1999:536-7, 2000:550-51).
Hirschman (1992) referred to three social mechanisms: exit, voice, and loyalty. Though all three can apply in varying ways to each sphere, exit refers primarily to the market sphere where, in a competitive situation, one has unlimited choice of buyers or sellers, so can “exit” from any one. Voice might refer primarily to the political sphere, where one can attempt to influence results by persuasion, and loyalty to the community sphere – though one could argue the other way as well.
Streeck and Smitter (1985:1) refer to these “three basic mechanisms of mediation or control” (Ouchi, 1980) as spontaneous solidarity, hierarchical control, and dispersed competition.
Friedland and Alford (1991:39) refer to three domains with different “logics of action”: In the marketplace, we are more likely to base our actions on individual utility and efficient means; in the polity, on democracy and justice; and in the family, on mutual support.
Van Staveren (2001:24) asserts that “three values appear time and again in economic analysis: liberty, justice, and care. Markets tend to express freedom, states to express justice, and unpaid labor to express care among human beings.” She notes (p. 213) that Ayres (1961:170) asserted a similar set of core human values: “freedom, equality, and security”. Van Staveren (p. 203) also notes:
- the form that these values take: exchange, redistribution, and giving;
- the locations where they operate: market, state, and the care-economy; and
- the corresponding virtues: prudence, propriety, and benevolence.
She further asserts that there are “distinct emotions and forms of deliberation as well”.
Mackey (2002:384) refers to “economic, political, and social problems” in Saddam’s Iraq; elsewhere (p. 181) she uses a different order, referring to “the new political, social, and economic paradigm” (an order which Rothstein and Stolle, 2007:1, also use); and yet elsewhere (p. 49) she notes that something “meant more socially, politically, and economically”. The order of expression doesn’t seem to matter, to Mackey or to most other authors, and one can easily find the other three permutations as well (e.g., Friedman, 2000:131; Giddens and Pierson, 1998:89; Sage, 2003).
But the community sphere is often ignored, and thus is sometimes considered third (Adaman and Madra, 2002). In political theory, the “Third Way” (Giddens, 1998) represents an alternative to either markets or governments, focused more in communities.
Waterman (1986:123) asserts “three freedoms: economic, political, and religious (conscience)”; and Hobson (1938/1976:52) refers to “the democratic triad of liberty, equality, fraternity”.
As some of these examples illustrate, a wide variety of words are used to refer to the three spheres, as in the title of the book (cited by Bennett, 1985) Mexico: Catholicism, Capitalism, and the State, or when
- Mackey (2002:217) discusses “political, economic, and… cultural control”;
- Bowles (1998:105) refers to “states, communities, and markets”;
- Wright (2000:211) refers to “governance, moral codes, and markets”;
- Mauss (1925/1967:52) refers to the “law, morality, and economy of the Latins” and to “the distinction between ritual, law, and economic interest”;
- Yuengert (1999:46) discusses “free markets circumscribed within a tight legal framework, and operating within a humane culture”;
- Polanyi (1997:140), in discussing “economic life”, refers to “freedom under law and custom, as laid down and amended when necessary by the State and public opinion”.
In The Foundations of Welfare Economics (1949:230), Little points out that “if a person argues that a certain change would increase economic welfare, it is open to anyone to argue that it would decrease spiritual or political welfare.”
This tripartite taxonomy has been used by economists since Adam Smith who, of course, had first written The Theory of Moral Sentiments (1759/1982) about communities and social goods, then The Wealth of Nations (1776/1976) about markets, economics. But he was planning a third major work – which was never completed – on the political system (Smith, 1759/1982:342 and “Advertisement” therein).
Minowitz (1993) uses the same tripartite taxonomy twice (in varying order) in the title of his book: Profits, Priests, and Princes: Adam Smith’s Emancipation of Economics from Politics and Religion.
The English economist and theologian Philip Wicksteed referred to “business, politics, and the pulpit” in his book of sermons titled Is Christianity Practical? (1885/1920, referenced in Steedman 1994:83). In discussing Wicksteed’s work, Steedman (p. 99) also refers to “potatoes, politics, and prayer”. Similarly, Hobson (1938/1976:55) referred to “the purse, power, and prestige of the ruling classes in business, politics, and society”. Success itself is often defined as “wealth, fame, and power” (Bogle, 2004:1; Carey, 2006), or sometimes as “money, status, and power”.
A similar tripartite taxonomy – perhaps Marxian – of firms, social classes, and states, can easily be seen as referring to the three spheres.
According to Trotsky (1957:255), communism would demonstrate that the human race had “ceased to crawl on all fours before God, kings, and capital” (quoted by Minowitz, 1993:240).
A variety of sources also provide evidence of an apparently widespread belief that the three spheres are both fundamental and exhaustive of social space. Michael Novak refers to the “three mutually autonomous institutions: the state, economic institutions, and cultural, religious institutions” as “the doctrine of the trinity in democratic capitalism” (Abdul-Rauf, 1986:175; also Neuhaus, 1986:517).
Dasgupta (1993:104) notes “one overarching idea, that of citizenship, with its three constituent spheres: the civil, the political, and the socio-economic.”
Meyer et al. (1992:12) assert that “individuals must acquire the means to participate effectively in the economic, social, and political life of the nation.” In the same work, Wong (1992:141) makes it clear that these three spheres are considered exhaustive by referring to “all social domains… economy… polity… and… cultural system”.
Polanyi (1997:158) describes the Russian Revolution and the Soviets’ “project for a new economic, political, and social system of mankind”.
Shadid (2001:3) points out that “political Islam, or Islamism…suggests an all-embracing approach to economics, politics, and social life.”
Dicken (2007:538) says that “corporate social responsibilities span the entire spectrum of relationships between firms [and] states, civil society, and markets.”