
Part Of: Demystifying Life sequence
Followup To: An Introduction To Natural Selection
Motivations
In the past two posts, we have explored the landscape of gene-space.
- A Genotype Is A Location.
- Organisms Are Unmoving Points
- Birth Is Point Creation, Death Is Point Erasure
- Genome Differences Are Distances
- Biological Fitness Is Height
Within this topography, we identified the following features:
- A Species Is A Cluster Of Points
- Species Are Vehicles
- Genetic Drift is Random Travel.
- Natural Selection is Uphill Locomotion
We can imagine millions of alien worlds with their own landscapes. Today, we survey the complexities of our biosphere: the fitness landscape of Earth.
Fitness-As-Resource
Let population be members of a species that are in the same geological area (such that its members are able to interbreed). For example, we know of fourteen different populations of the humpback whale. Now, for every population, there exists some maximum number of individuals that the local environment can sustain. This number is known as carrying capacity. Now, for most populations this number is fairly large, especially in fertile environments. What does this concept entail for our fitness landscape?
We can define fundamental fitness to be fitness that would be afforded to some pair of individual organisms in an empty world, with no other organisms competing for the same resources. As we introduce more organisms into a given population cluster, the amount of realized fitness we could afford the original members decreases. When the number of dots equals the carrying capacity, average fitness is 1.0 (such that population as a whole will neither increase nor decrease).
This perspective leads us to viewing realized fitness as a finite resource. This interpretation is entirely compatible with our physics-oriented view of life; life as disentropy engine. There is only so much disentropy to go around in a given volume of spacetime! Call this the fitness-as-resource view.
Biotopic Landscapes
Organisms successful in the jungle are not necessarily well-equipped for the desert. Fitness landscapes change with location. We must embed gene-space into spacetime.
Does every cubic centimeter of Earth’s surface merit its own gene-space? Surely not! Not even the most life-friendly milliliter cannot sustain thousands of lifeforms. We instead need to zoom out, and consider larger slices of land that can support a more meaningful amount of life. Will any collection of land work? No: we want to carve nature at the joints, and draw lines around ecologically uniform habitats (i.e., biotopes).
If we think of time as a dimension, then it is possible to view the entire universe stretched out throughout eternity as a four-dimensional block. But for our purposes, we only care about Earth across all time; call this smaller 4D rectangle the 4-biosphere. Ecological embedding is the art of finding maximally large 4-biotopes that maintain coherent landscapes..

Niches → Correlated Peaks
If organism fitness is a resource, what is resource competition? To answer, we must don our fitness-as-resource glasses. The idea here is that every organism comes equipped with disentropy vacuums; that is, machinery for extracting viable energy from its environment.
Let’s get specific. Imagine two different whale species in the same biotope, consuming the same kind of plankton. If the underlying plankton population started dying off, both whale populations would be jeopardized. Their disentropy vacuums would falter, and their collective fitness would plummet concurrently. Resource competition is fitness correlation.
Niches ultimately describe the resource-seeking strategies an organism uses to survive. Organisms with complete overlap of such strategies are said to occupy the same niche. While we could explore niches as a resource-space (the Hutchinsonian view, which has produced the discipline of niche modeling), let us instead view niches in terms of fitness landscapes. On this view, niches simply are correlated fitness peaks.
With this identification in hand, we are now in a position to understand more complete ecological phenomena:
- The competitive exclusion principle holds that, other things being equal, two species competing for the same resource cannot coexist at constant population values. In our language: housing multiple populations on the same correlated fitness archipellago is an unstable zero-sum game.
- Niche differentiation is a direct consequence of competitive exclusion. If another population invades and begins to take over your niche, you move to another niche (another resource profile). A classic example of niche differentiation comes from Robert MacArthur’s analyses of different populations of starlings in the same biotope. He observed that, despite their similar lifestyles, each population evolved to live in different cross-sections of trees: some living in the top branches, and others making homes near the base.
With niches under our belt, we now turn to two other, central notions in population ecology: food webs and arms races.
Food Webs → Lossy Fitness Theft
Our fitness-as-resource view provides a natural understanding of food webs. What is the fundamental fitness of the antelope? Zero! Herbivores cannot sustain themselves without vegetation; their existence is contingent on consuming such resources. Antelopes become relatively more reproductively successful only when grass becomes relatively less successful. This is a destructive form of fitness relocation, which I will call fitness theft.
93% of all human consumption of meat comes from three animals (36% pigs, 33% chicken, and 24% cows). None are carnivores. Why?
Consider the energy budget of life. If some blade of grass absorbs X kiloJoules worth of sunlight in its lifecycle, will the cow who ingests it absorb all X units of energy? No: most of the underlying energy was spent maintaining the cellular structure of the animal. Similar analyses can be run at any level of the food web. Life spends energy merely to sustain itself; this is the meaning of metabolism. Therefore, predation is always inefficient, and fitness theft is always lossy.
We can encode food webs into fitness landscape as follows. Diminishing peak size represents energy loss.
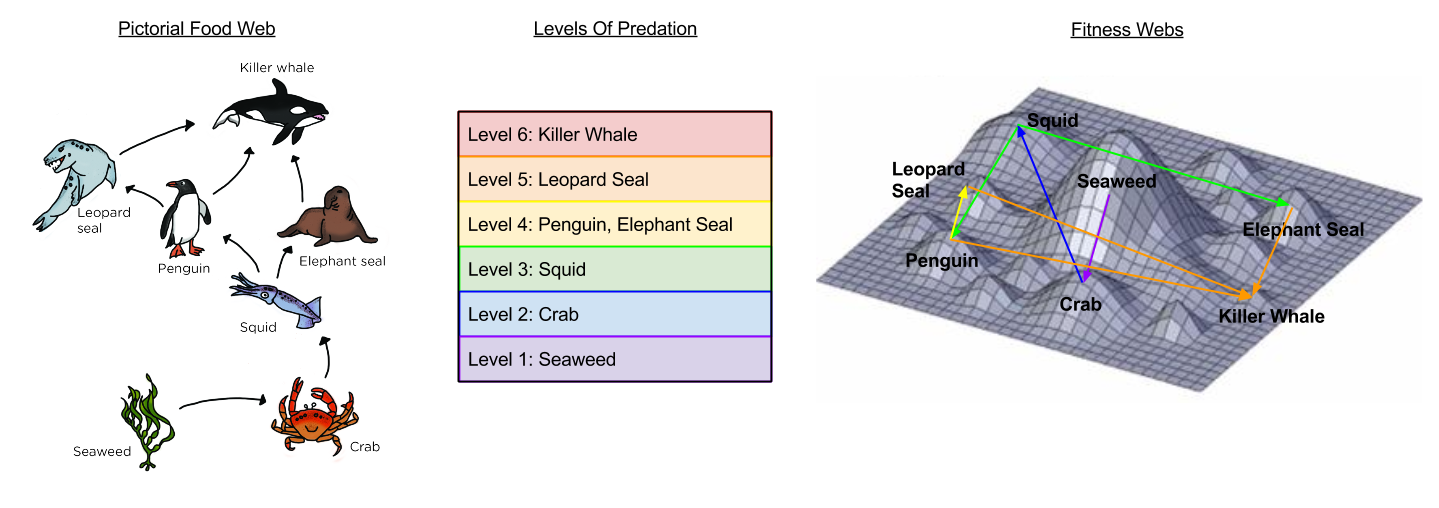
Perhaps unsurprisingly, food webs tend to create dynamical population patterns that fluctuate in sync with one another. Systems theory plays a role in their analysis; here is a visual guide to the underlying differential equations.
Arms Race → Comoving Peaks
Meet the American rough-skinned newt.

Cute, right? Just, if you hold one, please remember to wash your hands before eating. These things secretes enough poison to literally kill an elephant.
Why should this be? I mean, obviously poison is a good safeguard against predation, but none of the newt’s natural predators come close to being elephant-sized. What benefit could the newt possibly derive in developing a poison so gratuitous? Genetic accidents seems unlikely to explain the immense gap between the practical dose and the actual dose. What gives?
The clue lies in looking carefully at the newt’s predators. It turns out that a nearby species of garter snake has been developing a massively overpowered immunity to the newt’s poison.
If predators can be viewed as “fitness vacuums” in gene-space, then predation can induce natural selection towards novel defense mechanisms. As prey evolve towards more defensible peaks, predators with bolstered offensive capabilities are selected. In this way, the peaks of both species move alongside one another:
Takeaways
As hinted in my reference to systems theory above, theoretical ecology does not always leverage fitness landscape models.
We began this post by naturalizing our notion of gene-space:
- Fitness is ultimately grounded in energy budgets. Fitness is thus a finite, fungible resource.
- The fitness landscapes of a desert diverges from that of the ocean. Fitness landscapes are most clearly defined in uniform habitats, or biotopes.
These theoretical additions let us model new ecological behavior:
- Niches are correlated fitness peaks, where each peak “vacuums up” fitness from the same set of resource.
- Food webs are fitness theft, where predators gain fitness by reducing fitness of their prey. However, predation is inefficient, which guarantees a finite number of predation levels.
- Arms races are comoving peaks, which occur when predator and prey attempt to outmaneuver one another in the fitness landscape.
These phenomena suggest that the fitness landscape is better understood as a seascape, whose contours fluctuate & interrelate in subtle ways.

{kind=link}