
Part Of: Anthropogeny sequence.
Content Summary: 1600 words, 16 min read
For all his noble qualities, godlike intellect, and exalted powers, man still bears in his bodily frame the indelible stamp of his lowly origin.
– Charles Darwin, Descent of Man
Setting The Stage
Common descent denotes the discovery that all species are related: that living organisms reside in a single tree of life. Homo Sapiens is no exception. We diverged from other hominoids (great apes) some 7 mya. During that time period, fossils more than 6,000 individuals from dozens of bipedal ape species.

Today, we explore why apes became bipedal. But first, the evolution of apes.
Primate Evolution
Primates are mammals with flat nails instead of claws, grasping hands and feet, a highly developed visual system. They are highly iteroparous (long juvenile period) and have large brains to support the complex needs of group living. Primates are known for their symbolic dominance hierarchy, friendship mediated by grooming and mindreading (making inferences about the mental state of their peers).
Apes are primates that hang from branches (no tail), and even larger brains that promote behavioral flexibilities. Apes are known for coalitional warfare, group-specific cultural behaviors, flexible group signaling (e.g., mobbing), and tool-making.
The primate lineage emerged in the Paleocene (60 mya); apes in the Miocene (20 mya).

Without a tail (and in a “dead-end” body plan that precludes growing it back), apes increasingly relied upon behavioral flexibility to mitigate their comparative immobility. A monkey is an ecological specialist; the ape lineage was populated by generalists.
Apes flourished in the early and middle Miocene (20-10 mya). But they began to die out, starting in the late Miocene (10 mya). Today, there are hundreds of extant species of monkeys, and only five apes (gibbons, gorillas, orangutans, chimps and bonobos).
Evolution and progress are not synonymous. The ape branch of the tree of life is sparse because we are a failed lineage.
The failure of our ancestors seems to have been driven by a radiation from earlier primates (monkeys) in what can be called revenge of the specialized. It became increasingly difficult for generalized omnivorous species to find niches that were not more effectively exploited by a whole host of small-sized specialist monkeys.
Amidst this harsh inter-primate competition, it is interesting to note that modern apes are substantially larger than their Miocene ancestors. An increase in the body size of living apes and humans may well represent an evolutionary response to competition from monkeys.
We turn now to the question of bipedality. Before we can address why apes stood on two legs, we must first understand the anatomy of bipedality.
The Anatomy of Walking
The main anatomical structure that changed was the pelvis. The pelvis is not a single bone, but rather three bones glued together by cartilage. As we will see shortly, bipedality requires shortening of the ilium.

Walking is a pendulum-like motion. Most of the time one foot is off of the ground. This provides a stabilization problem. To solve this, bipedal animals have abductor muscles. You can actually feel these yourself: next time you walk around, feel the muscle on your hips flex (but only the muscle on the side of the weighted foot).

Abductor muscles aren’t enough, however. In order to further stabilize a two-legged gait, the legs must be brought closer together. Adjusting the femur angle brings the center of gravity closer together:

Finally, to improve the energy efficiency of walking, the human foot transitioned from a grasping surface to an energy-transfer platform.

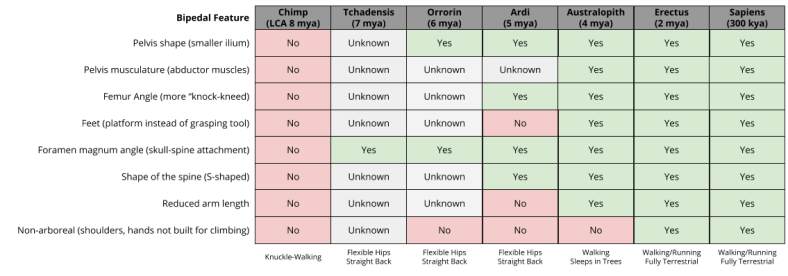
We have so far discussed four features of bipedal living. Here is a more complete list:
- pelvis shape (smaller ilium)
- pelvis musculature (abductor muscles)
- femur angle (more “knock-kneed”)
- feet (platform instead of grasping tool)
- foramen magnum angle (how the skull attaches to the spine),
- shape of the spine (bipedal spines are S-shaped), and
- reduced arm length (no longer needed to contact the ground)
The definition of hominin is bipedal ape. Little surprise then, that even the earliest hominin (Sahelanthropus Tchadensis) has at least one feature associated with bipedalism. As we move to more recent species, we can see increasingly “classical” body plans:
Bipedality also explains why human beings suffer from:
- Lower back pain. For hundreds millions of years, the spine was housed on a horizontal chassis. Switching to a vertical chassis places a lot of pressure on the lower spine. Zebras don’t suffer from lower back pain as much as human beings.
- Hernias. The strain is not limited to the skeleton. Pressure also dramatically increases in the lower abdomen, causing an unusually high rates of hernias for human beings. In fact, one of the distinguishing characteristics of human beings is our smooth, fatty skin. We preferentially store fat subcutaneously to combat the pressure in our abdomen.
Theories of Bipedality
The fact that African apes became bipedal around 6 mya is not particularly interesting. A more interesting question is why African apes became bipedal. How did bipedality amplify the hominin niche?
There is no shortage of theories. Here are six:
- Brachiation (arm-based locomotion via branch-swinging) responsible for the postcranial features we share with apes.
- Arboreal apes modified their vertical climbing to walk bipedally along thick branches in the canopy.
- Bipedalism emerged from the need to carry babies, food, and other objects back to base.
- An aquatic phase of foraging and avoiding predators in water.
- Predator avoidance in the savannah with frequent peering over tall grass.
- A thermal theory whereby savanna dwellers stand up to keep cool.
These theories leave much to be desired, however.
First, some disregard ecological data entirely. The last two theories rely on the savannah hypothesis: that standing on two legs was made advantageous as forests increasingly disappeared. But the savannah hypothesis is wrong. Bipedalism emerged 6 mya, but the savannah grasslands only appeared 2-4 mya.
Second, they disregard the incrementality of natural selection. Two-legged standing preceded true bipedal walking and should not be lumped with it. We must conceive of an ape that can stand but not walk (Orrorin tugensis?), and an ape that can walk but not run (Australopithicus afarensis).
More generally, whenever we see a complex adaptive package like walking, it is immediately useful to explore prerequisite abilities. One natural way to conceptualize the increments is as follows:
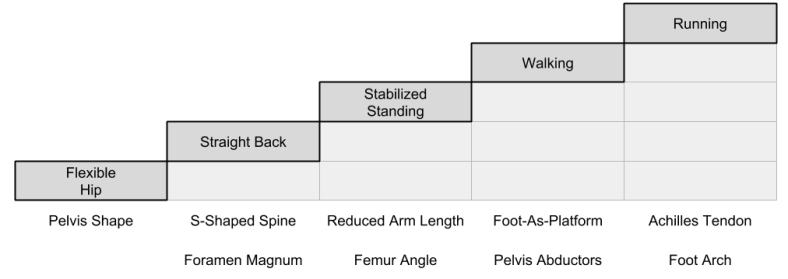
The above image identify anatomical increments with each new behavioral capability.
We are not looking for a single ecological incentive for bipedalism; rather, we need individual motives for each increment in the journey to bipedality.
What kind of niche would reward flexible hips and a straight back?
The Primacy of Ecology
To answer this question, we need to get familiar with African geology and ecology.
As the most common promoter of diversity, allopatric speciation occurs when some population becomes isolated from the broader gene pool. Typically, these episodes are caused by climate change: the species gets “locked in” to a particular area by encroaching deserts, and then expands to surrounding habitats once the desert recedes.
The African continent contains wet-spots (equational rain) and hotspots (deserts). During cold glacial periods, these wet-spots expand along an east-west axis. For warm interglacial periods, the hot-spots expand along a north-south axis.

There are two primary forests in Africa:

During the most arid climatic phases, the desert corridor separating these forests would close, leading to genetic isolation and speciation.
Squat Feeding in the Eastern Littorals
What kind of ape would emerge from the Main Forest Block? Such species would remain conservative (change slowly) because their much larger range embraces a much wider range of different types of wooded habitats. In fact, we know that modern-day gorillas derive from this ecosystem.
What kind of ape would be forged by the Eastern Forest Littoral? This smaller, fragmented ecosystem would cause both selection and genetic drift to accelerate. There are several peculiarities to this ecosystem worth pointing out:
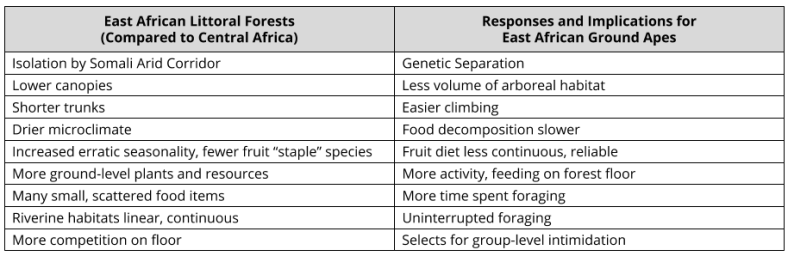
In short, apes isolated in East African littoral forests seem likely to have found a niche on the forest floor. The natural distribution of resources favors this interpretation; and the growing competition from monkeys would have made the canopy increasingly infeasible.
These ground apes faced strong selective pressure to improve their foraging efficiency. The chimp pelvis has a very long ilium, which “locks into” the ribcage. There are clear foraging benefits for a reduction in the ilium (flexible waist), and straightening of the back (improved visibility).
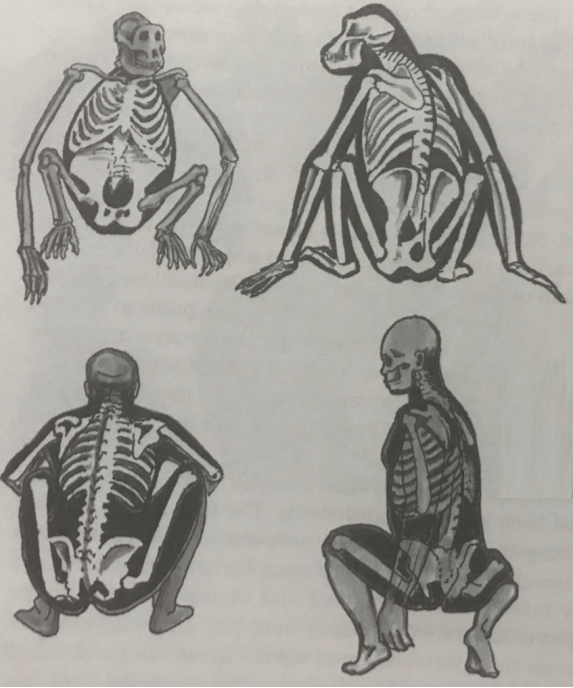
In short, the squat-feeding hypothesis explains why flexible hips and straight spines were selected in ground apes of the early Pleistocene.
Other adaptive explanations only become relevant in further increments of the transition to bipedality. In particular, starting around 4 mya, the African continent began to dry. This made fruit increasingly less concentrated, and more seasonal. Locomotion thus became increasingly necessary to get enough calories.
In modern humans, walking is four times more efficient than chimpanzee knuckle walking. Of course, very ancient hominins like Ardipithecus Ramidus could walk, but were less efficient than the Australopiths (and us, for that matter). But clumsy walking merely needs to improve upon the kinematic efficiency of knuckle walking, which as we have seen is not hard to do.
Bipedalism is not universally advantageous. Hominins like us are half as fast as other apes, and we have lost the ability to gallop. Greatly reduced ability to change direction while running. The earliest bipeds probably avoided open habitats because of their increased vulnerability to predation, preferring forest and riverine habitats instead.

The facilitation of walking and running was not the ecological reason why our ancestors began the journey towards bipedality. But once they started on this particular anatomical pathway, these applications became possible. Thus, it is only with hindsight that we can say that the ultimate worth of standing up, the hidden evolutionary prize, was the ability to find the way out of a sort of ecological cul-de-sac.
Concluding Thoughts
The squat feeding theory of bipedality, as well as several of the images of this post, are credited to Jonathon Kingdon, African zoologist and author of Lowly Origin. I highly recommend this text, for those curious to learn more.
Until next time.
