
Part Of: Affective Neuroscience sequence
See Also: The Construction of Body Status
Content Summary: 3000 words, 15 min read
Physiological Control
What is life? Schodinger (1944) argued that life is a negentropy phenomenon: free energy is concentrated locally, in the face of surrounding energy dissipation. This explains several facts of biology:
- Inference. Jaynes (1957) famously argued that both statistical and biological inference derive from free energy minimization.
- Abiogenesis. England (2013) argues that replication can be understood in thermodynamic terms, and view abiogenesis as driven by negentropy.
- Metabolism. The field of bioenergetics (Cheetham 2010) has discovered metabolism to be consistent with this thermodynamical principle.
Homeostasis was described by Bernard (1878) as, “all vital mechanisms, however varied they may be, have only one object, that of preserving constant the conditions of life in the internal environment.” It has a thermodynamic interpretation: life is buffeting its internal milieu from the dissipative vagaries of the environment.
For example, blood sugar concentration is a biologically defended variable. Too much glucose causes energy poisoning, too little causes cellular starvation. The pancreas regulates blood sugar levels between 70 and 140 mg/dl. When blood sugar is too low, the pancreas releases glucagon to raise serum glucose; when too high, the pancreas releases insulin to reduce the level.
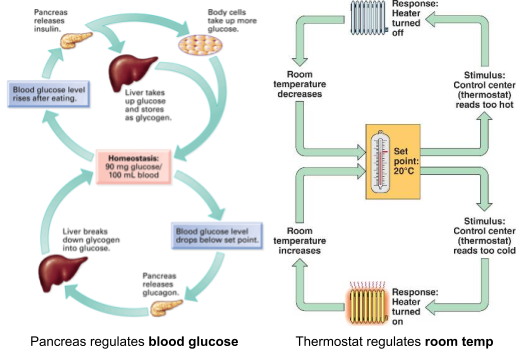
The pancreas here plays a similar role as a thermostat which is engineered to regulate room temperature. Thermostat design is heavily influenced by engineering control theory. The set point (i.e., the desired value) is specified, and the error term (difference between observation & set point) is computed. The error term can be used in two different ways:
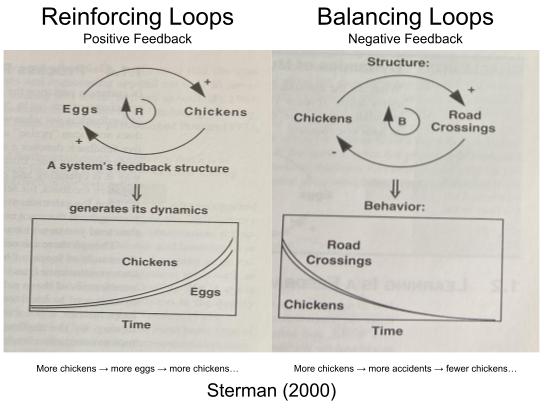
- Positive feedback tends to produce exponential growth. Evolutionary arms races are a biological example, microphone squealing is an electronic example.
- Negative feedback tends to stabilize a system. Physiological regulation may be a useful example…
Feedback loops are a central organizing principle of business dynamics (i.e., systems thinking). According to Sterman (2000): “all systems, no matter how complex, consist of overlapping networks of positive and negative feedbacks, and all dynamics arise from the interaction of these loops with one another.
The brain regulates body adiposity via the lipostat (Cabanac & Richard 1996), largely mediated by the leptin system (Zhang et al 1994).
- After overfeeding (Sims et al 1968), the body will attempt to lose weight (more metabolism & less food intake).
- After underfeeding (Key et al 1950), the body engages a starvation response to gain weight (less metabolism & more food intake)
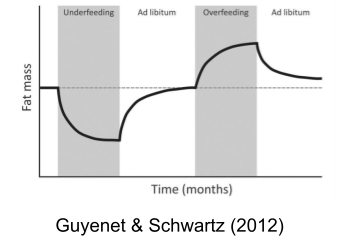
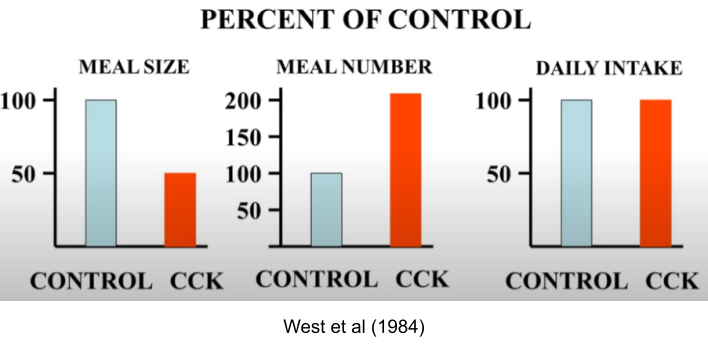
Taken together, these effectors work to maintain a constant level of stored calories (adiposity). Adiposity is biologically defended, which is why willpower-based dieting interventions so often fall short. If you inject a satiety hormone cholecystokinin (CCK) into a rat, meal size roughly halves.. but the number of meals initiated doubles to compensate.
The accuracy of your body’s regulatory systems is remarkable. For eating, annual energy intake is 955,570 calories; gaining one pound is 4,000 calories. In other words, your lipostat adjusts your energy intake to equal energy output with an error of 0.4%, or 11 calories per day. That’s a potato chip.
Adjustable Set Points
When Cannon (1929) first introduced homeostasis, he emphasized set point constancy. Yet set points can change; these are called rheostatic adjustments (Mrosovsky 1990). Consider body temperature. When a rat encounters an overly hot room (a forced change), it will compensate via three cooling effectors: skin blood flow, evaporation, and behavior to find a cooler environment.
Body temperature can also become elevated in response to infection, which helps the immune system combat pathogens. But this regulated change is handled differently by the body; with fever, warming effectors are engaged: metabolism increase (shivering), piloerection (goosebumps), and behavioral motivation to find a warmer environment. Only in fever is the set point adjusted.
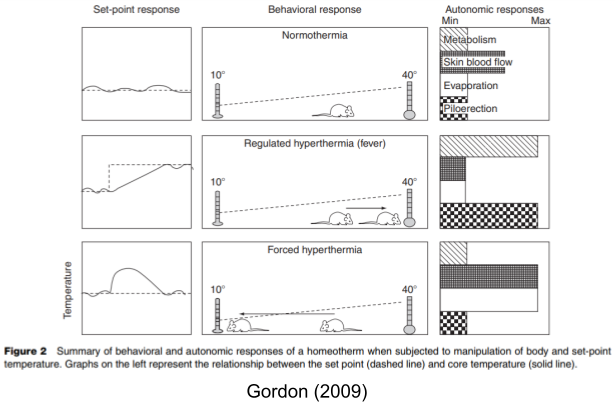
Other set points change too. The blood pressure set point is also dynamically altered from normal waking (~85 mmHg), sleep (~60 mmHg) and periods of extended stress (~120 mmHg).
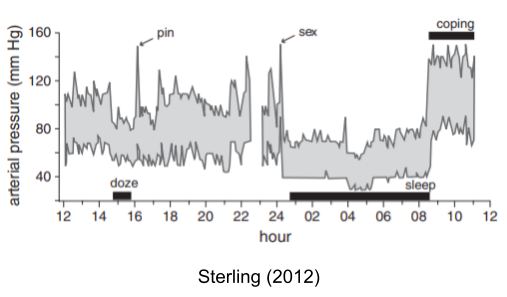
Many other example of circadian rheostatic adjustments exist:
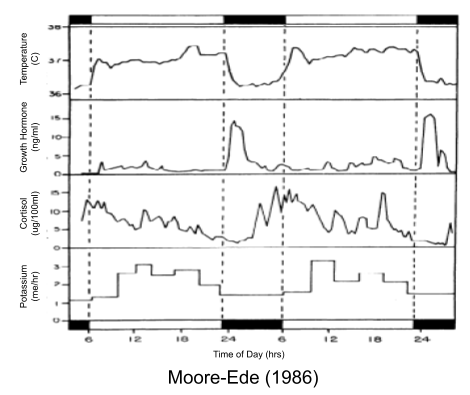
Why? What is the relationship between physiology and chronobiology?
Reactive vs Predictive Homeostasis
Homeostasis is a central organizing principle of the viscera. But chronobiological rhythms is another such principle. Scientists once believed that such rhythms were direct responses to zeitgebers (environmental time cues). In 1729, a French astronomer placed a plant in a dark closet where it was no longer subject to daily variations in illumination. To his surprise, the plant continued to raise and lower its leaves on schedule. De Mairan (1729) concluded the plant had an internal clock for telling the time. This clock endogeneity result was shelved for a century, but was ultimately replicated.
Why should the nervous system mold behavior into such predictable rhythms? As Moore-Ede (1986) puts it,
Amidst the random variations in environmental opportunities and challenges, two highly regular geophysical cycles – the year and the day – stand out. They dominate multiple aspects of our environment, most directly illumination and temperature. One of the most important features of these environmental cycles is extreme predictability. The time it takes for the rising sun to return to shine on the same spot on the spinning Earth (the solar day) varies by no more than a few minutes in any 1 year, and the average period of the Earth’s daily rotation has only slowed by 20 seconds during the course of the last million years. Since these cyclic changes are so predictable, it is scarcely surprising that circadian and circannual adaptive mechanisms have evolved to take advantage of this predictability.
Circadian rhymes manifest in sleep-wake behavior, feeding-fasting cycles, drinking behavior, melatonin synthesis, and locomotor activity. In the controlled atmosphere of the lab, they rarely change; but in natural settings they do. Their phase flexibly responds to changing energetic conditions: a kind of temporal niche switching (Riede et al 2017).
Lesions of the suprachiasmatic nucleus (SCN) in the hypothalamus abolish these rhythms (e.g., Eastman et al 1984). Here, for example, is drinking record of a squirrel monkey – SCN lesions do not change the overall volume intake, but it does result in arrhythmia.

Consider a rat food dispenser that dispenses calories only during a 3h window. The rat will ramp up its food seeking behavior (lever presses) about an hour before the food is available. If the food availability window is shifted forward by six hours, the predictive behavior occurs on schedule, but the reactive behavior only occurred when the press of the lever first produced a pellet at 2pm. Over the course of the next eight days, the predictive component gradually slid towards the new window.
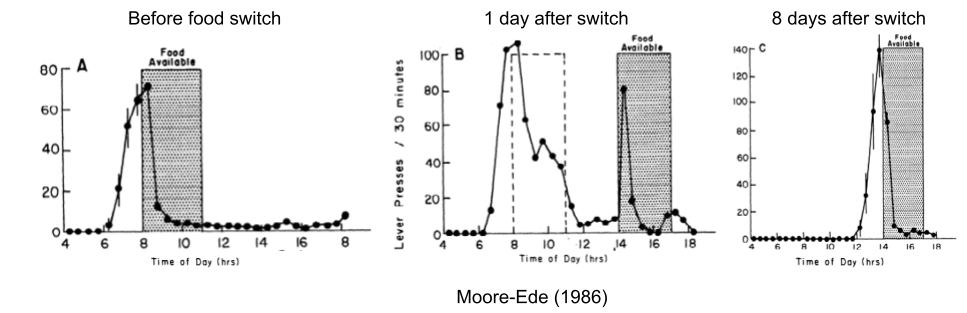
If a rat becomes motivated to eat only when it is hungry, that is too late. Foraging is a time consuming behavior. And the body itself needs time to prepare for a meal: blood glucose decreases before a meal, which serves to blunt the postprandial rise in glucose. These anticipatory cephalic phase responses (CPRs) enable animals to cope with the imbalance created when food is restored.
Decoupling of predictive and reactive components has been replicated in myriad other domains (Moore-Ede 1986, Schulkin & Sterling 2019). In general, phase adjustments occur when the original rhythms fail (Riede et al 2017). Errors in predictive homeostasis promote exploration of alternative temporal niches.
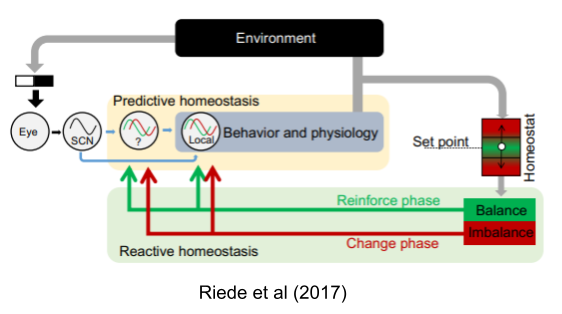
Finally, predictive homeostasis is not limited to circadian clocks. Arbitrary cues can elicit homeostatic responses.
Can the concept of homeostasis include predictive, error preventive components. Researchers disagree. Moore-Ede (1986) says yes, Schulkin & Sterling (2019) says no and reserves the term allostasis for the latter concept. Much of this dispute is taxonomic, and centers on interpreting Cannon (1929)’s original meaning. In the final analysis, however, both sides have converged on a dual process theory of regulation.
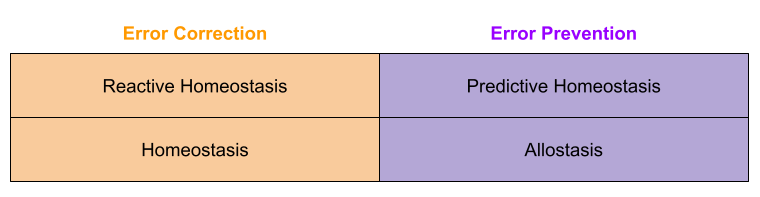
Balance Point Theory
The concept of set point may need to be discarded. To understand why, let’s return to thermoregulation.
In humans, we think of a single set point of 98.6 F. But your CNS measures two quantities: peripheral temperature (e.g., in the skin) and core temperature (e.g., in the stomach). Peripheral temperature sensors are not used exclusively by thermoregulation, they also push fine-grained spatial data to the insula for more specific interactions with the environment (e.g., burn prevention).
Due to thermal inertia, the core changes more slowly than the periphery (especially in large animals). This makes the periphery a leading variable. Because skin is more predictive than core, its sensors are more likely to evoke a preventative behavioral response (Romanovsky 2007).
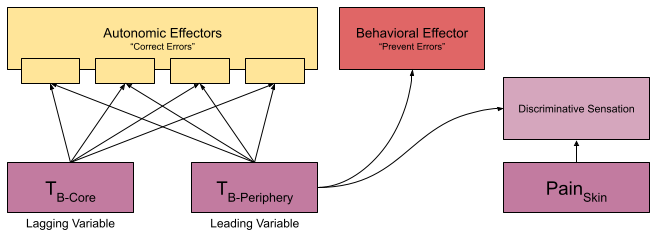
Some autonomic effectors are more responsive to core temperature and vice versa. McAllen et al (2009) report experiments where they immersed a rat in water & rapidly altered the skin temperature. By measuring effector neurons during this experience, they showed that these effectors do not respond univocally – they differ in responsiveness to changes in core and skin.
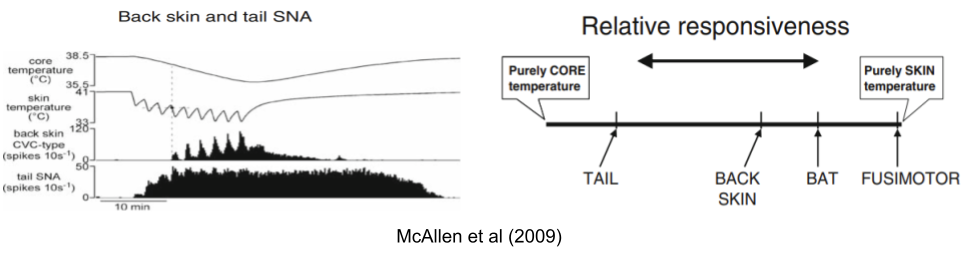
These effectors not only respond to different inputs. Each input responds to different temperatures, and each output flows through an independent anatomical pathway:
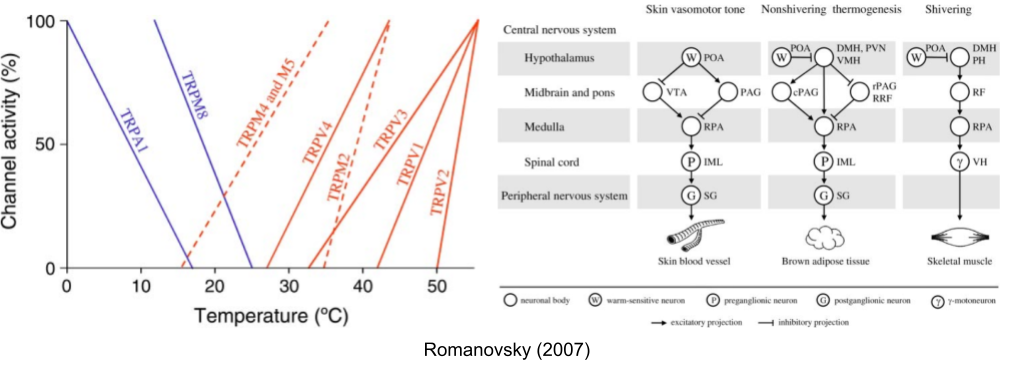
The classical theory of homeostatic set point requires a central controller, which aggregates across temperature sensors to compute a mean temperature & coordinate its responses. But the anatomical basis for such a system remains elusive. And we have seen that thermoregulation effectors have independent inputs, functions, and outputs.
Satinoff (1978) proposes that the central controller does not exist, and neither does a single set point. She modeled multiple, independent sensor-effector loops, whose arithmetic sum of activity across activated effectors comprise the biologically defended range. As the subpopulation of active sensor-effector loops changes, so too does the aggregate balance point (Romanovsky 2007).
Decentralized control systems envisioned by balance point theory are self-organizing. They have important advantages, including flexibility and robustness to loss of system components (Seeley 2002). This comes at a price: decentralized systems cannot guarantee optimality, in part because of inefficient coordination of the distributed subsystems. Decentralized control may explain the following phenomena:
- Opponent Processes. Simultaneous use of the furnace and air conditioner wastes fuel and contributes to wear and tear on both pieces of equipment. In the same way, it is hard to imagine a central controller producing such an inefficient result, but balance point theory explains the phenomenon quite nicely. One popular model of addiction assumes that reward and hedonia are physiologically regulated and that sensor-effector loops exist that influence the value of these variables (Koob & Le Moal 2008). Addiction is viewed as competition between a generic antireward mechanism versus a specific cue being ascribed increasing amounts of incentive salience.
- Asymmetric Enforcement. For adiposity, mammals are very efficient at responding to underfeeding (and surgical removal of fat), but their response to overfeeding (or surgical implantation of fat) is much less robust. Leptin in particular seems to enforce the lower threshold alone (Wade 2004). For temperature, mammals exposed to severe stressors (e.g., endotoxins) will shut down thermogenesis and revert to poikilothermy – the upper threshold is still defended, but the lower defended threshold drops by tens of degrees (Romanovsky 2004).
- Overcompensation. Cold-challenged armadillos resulted in persistent and excessive increases in core temperature and oxygen consumption that exceeded the pre-perturbation values (discussed in Ramsay & Woods 2014). Similarly, Slot et al (2002) show that exposure to fentanyl at first produces pain tolerance, but over time leads to pain intolerance (hyperalgesia):
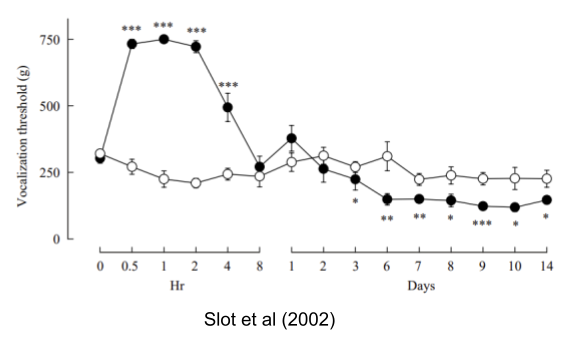
Overcompensation may be involved in many modern-day examples of cumulative movement of the balance point (aka rheostatic drift), including
- why BMI began increasing in the 1970s (James et al 2001);
- why sperm count is declining (Levine et al 2017);
- why body temperature is falling (Protsiv et al 2020);
- why autoimmune diseases are on the rise (Brady 2012); and
- why the placebo effect is strengthening (Tuttle et al 2015).
Market-based control (Clearwater 1996) represents one way to formalize balance point theory. Consider blood pressure. The body must balances between demand (organs requiring nutrient-rich blood) and supply (the heart-lung system which distributes these resources). Blood pressure can be interpreted as a price equilibrating supply and demand: higher blood pressure benefits organs but places a strain on the heart, and vice versa (Fink 2005). This interpretation of blood pressure casts new light on the strong associations between the hypertension, obesity, and hyperglycemia (i.e., metabolic syndrome): they are equilibria of an energy surplus.
Interloop Effectors
In Two Cybernetic Loops, we saw how the nervous system can be dichotomized as a world-oriented cold loop and body-oriented hot loop. Our discussion of homeostasis has centered on internal bodily variables being regulated by internal effectors. But one of the key functions of the hot loop is to “cross state lines” and motivate foraging behavior (i.e., external effectors) in service of the body.

The relationship between hot loop & reward system is complicated to tease apart. Activity in agouti-related protein (AgRP) neurons, aka hunger neurons, cause sensations of hunger, and also transmit a sustained positive valence signal that conditions both Pavlovian and instrumental learning (Chen et al 2016). When an animal sees food, AgRP neurons are immediately silenced, but the reward system nevertheless promotes consummatory behavior.
In obesity, we know that ultra-processed foods (UPF) play a role (Hall et al 2019). But we still don’t know which system is to blame! Is the epidemic caused by toxins (e.g Alharbi et al 2018) interfering with our internal effectors? Or by superstimulus (hyperpalatable foods) overwhelming our reward system (Guyenet 2017)? The jury is out.
Panksepp (1998) describes the basal ganglia as the SEEKING system, a generic mechanism that promotes both appetitive and consummatory motor behaviors. Activation of this system generates emotional experiences of curiosity, engagement, wanting, and motivation. Homeostatic detectors in the medial hypothalamus activate SEEKING via the lateral hypothalamus:
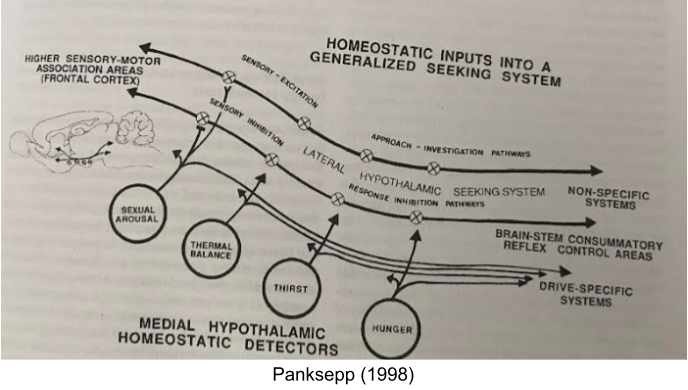
The reward value of a stimulus increases with the effectiveness of that stimulus in restoring bodily equilibrium (homeostasis). This effect, known as alliesthesia, is well-documented for food rewards, which are more pleasurable when they relieve a hunger state (Burnett et al 2019). Further, on different visceral states (e.g., hungry vs thirsty), the reward topography will change, despite the absence of any phasic DA learning signals.
Wanting is the substrate of arousal, or motivation. Its purpose is to control metabolic expenditure. We can see evidence for this in adjunctive behaviors. If a starving rat is given some food, but not enough to mitigate the drive, it will engage in ritualistic behaviors, such as pacing, gnawing wood, or running excessively. This phenomenon of behavior substitution was also discovered by ethologists under the heading of displacement behaviors. For example, two skylarks in combat might suddenly cease fighting and peck at the ground with feeding movements. These behaviors rely on the mesolimbic dopamine system (Robbins & Koob 1980).
Formal models (such as reinforcement learning and active inference) aspire to model the behavioral profile of the SEEKING system. Yet many models do not include these visceral influences. I look forward to the day a simulated agent can reproduce these phenomena.

Until next time.
References
- Alharbi et al (2018). Health and environmental effects of persistent organic pollutants
- Bernard (1878). Les phenomenes de la vie
- Brady (2012). Autoimmune Disease: A Modern Epidemic?
- Burnett et al (2019). Need-based prioritization of behavior
- Cabanac & Richard (1996). The nature of the ponderostat: Hervey’s hypothesis revived
- Cannon (1929). Organization for physiological homeostasis.
- Cheetham (2010) Introducing biological energetics
- Chen et al (2016). Hunger neurons drive feeding through a sustained, positive reinforcement signal.
- Clearwater (1996). Market-based control: a paradigm for distributed resource allocation.
- De Mairan (1729). Observation botanique.
- England (2013). Statistical physics of self-replication
- Falk (1970). The nature and determinants of adjunctive behavior
- Fink (2005). Hypothesis: the systemic circulation as a regulated free-market economy. A new approach for understanding the long-term control of blood pressure
- Gordon (2009). Autonomic Nervous System: Central Thermoregulatory Control
- Guyenet (2017). The Hungry Brain: Outsmarting the Instincts That Make Us Overeat
- Guyenet & Schwartz (2012). Regulation of Food Intake, Energy Balance, and Body Fat Mass: Implications for the Pathogenesis and Treatment of Obesity
- Eastman et al (1984). Suprachiasmatic nuclei lesions eliminate circadian temperature and sleep rhythms in the rat
- Hall et al (2019). Ultra-Processed Diets Cause Excess Calorie Intake and Weight Gain: An Inpatient Randomized Controlled Trial of Ad Libitum Food Intake
- James et al (2001). The worldwide obesity epidemic
- Jaynes (1957). Information theory and statistical mechanics
- Key et al (1950). The biology of human starvation.
- Koob & Le Moal (2008). Addiction and the brain antireward system.
- Levine et al (2017). Temporal trends in sperm count: a systematic review and meta-regression analysis
- McAllen et al (2010). Multiple thermoregulatory effectors with independent central controls.
- Modell et al (2015). A physiologist’s view of homeostasis
- Moore-Ede (1986). Physiology of the circadian timing system: predictive versus reactive homeostasis
- Mrosovsky (1990). Rheostasis: the physiology of change
- Panksepp (1998). Affective Neuroscience
- Protsiv et al (2020). Decreasing human body temperature in the United States since the Industrial Revolution
- Romanovsky (2007). Thermoregulation: some concepts have changed. Functional architecture of the thermoregulatory system.
- Romanovsky (2004). Do fever and anapyrexia exist? Analysis of set point-based definitions
- Ramsay & Woods (2014). Clarifying the roles of homeostasis and allostasis in physiological regulation
- Robbins & Koob (1980). Selective disruption of displacement behaviour by lesions of the mesolimbic dopamine system
- Satinoff (1978). Neural organization and the evolution of thermal regulation in mammals.
- Schulkin & Sterling (2019). Allostasis: a brain-centered, predictive mode of physiological regulation
- Schrodinger (1944). What is life?
- Seeley (2002) When is self-organization used in biological systems?
- Sims et al (1968). Experimental Obesity in Man.
- Sterman (2000). Business Dynamics
- Sterling (2012) Allostasis: a model of predictive regulation
- Slot et al (2002). Sign-reversal during Persistent Activation in l-Opioid Signal Transduction
- Tuttle et al (2015). Increasing placebo responses over time in U.S. clinical trials of neuropathic pain
- Wade (2004) Regulation of body fat content?
- West et al 91994). Cholecystokinin persistently suppresses meal size but not food intake in free-feeding rats
- Wing & Phelan (2005). Long-term weight loss maintenance
- Zhang et al (1994). Positional Cloning of the Mouse Obese Gene and its Human Homologue
