
Part Of: Neuroeconomics sequence
Content Summary: 1000 words, 5 min read
Reentrant Loops
The thalamus receives input from the entire cortex. The cortex & thalamus like two nested spheres, innervating one another. Let us call such cortical-subcortical loops, reentrant loops.
The basal ganglia also participates in a reentrant loop. The basal ganglia and thalamus are nestled inside one another, far below the cortical mantle:

The last major reentrant loop involves the cerebellum. Taken together, reentrant loop anatomy looks something like this:

Dopamine Anatomy
We turn now to the basal ganglia. The basal ganglia is innervated with dopamine, and contains an order of magnitude more dopamine receptors than any other brain region (Dawson et al, 1986). To understand this neurotransmitter, we must consider its biochemical signature.
As a molecule, dopamine contains catechol (a kind of benzene C6H6) and a side-chain amine. Thus, it participates in the catecholamine family.
There are in fact many kinds of catecholamines. For example, from phenethylamine you can synthesize tyrosine (via hydroxylation). In fact, there are thirteen different catecholamines, with a particular tree of biosynthetic sequences:
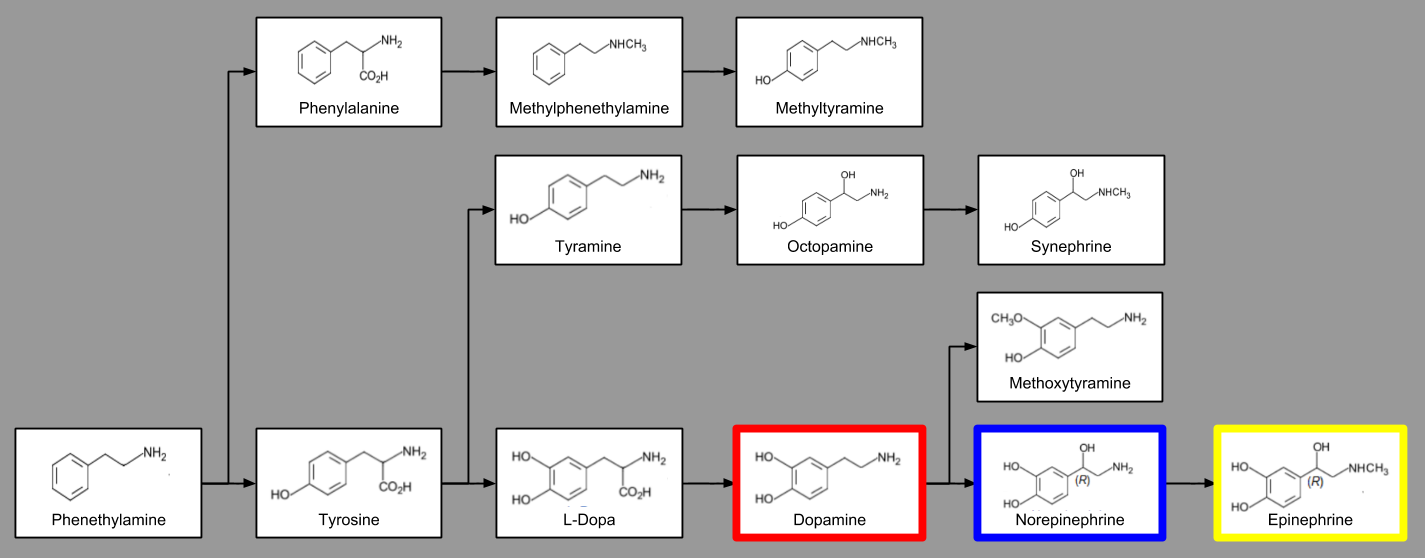
Three particular catecholamines are major players in brain neurochemistry.
- Epinephrine (EPI), i.e. adrenaline, promotes the fight-or-flight response.
- Norepinephrine (NE) encodes perceptual salience.
- Dopamine (DA) is the currency of motivation and reward.
Thus, catecholamines play a role in metabolic, perceptual, and motor arousal. In short, they are arousal systems.
Within the brain, a small number of discrete cell groups manufacture dopamine:

In the above cell groups, two dopaminergic cell groups are particularly important: A9 (SNc) and A10 (VTA). The nigrostriatal pathway emerges from the former, and the mesolimbic and mesocortical pathways emerge from the latter.

Basal Ganglia Anatomy
The basal ganglia is an ancient structure, with four main components:
- The primary input structure, the Striatum
- Global Pallidus External (GPe)
- Subthalamic Nucleus (STN); and
- three interrelated structures: the Global Pallidus Internal (GPi), Substantia Nigra Pars Reticulum (SNr), and Ventral Pallidum (VP)
These components comprise the core of the CBTC Loop:

In the above, basal ganglia nodes are in purple. Black arrows are excitatory, red are inhibitory, green are dopaminergic.
Three Circuits: Direct, Indirect, Hyperdirect
Can you find the loop in this circuit? In fact, if you look closely, there are three:

Consider the direct pathway. We see that the thalamus has an excitatory influence on the cortex. However, the GPi/SNr/VP node inhibits the thalamus. However, if an activated striatum inhibits the GPi/SNr/VP, the overall effect is “releasing the brakes”. The direct pathway is excitatory.
In the indirect pathway, the STN buffers the GPi/Snr/VP “stop” signals, and attempts to “release the brakes” from the GPe is inhibited by the Striatum. Three inhibitory tracts along the indirect pathway are net-negative. Thus, the indirect pathway is inhibitive.
The hyperdirect pathway has one inhibitory track (the GPi/SNr/VP “brakes”) – thus this pathway inhibits action. Why did evolution select two inhibitory pathways? One answer might rely on circuit length. The hyperdirect pathway bypasses several stages of the basal ganglia: perhaps its use was to accelerate the inhibitory signal of the indirect loop.
The most prominent cell type in the striatum is the medium spiny neuron (MSN). MSNs express one of two kinds of dopamine receptors: D1 and D2. Cells expressing these receptors are thoroughly mixed in the striatum. However, these pathways are strictly segregated by dopamine receptor:
- D1-expressing MSNs only participate in the direct pathway, and
- D2-expressing MSNs only participate in the indirect pathway.
Basal Ganglia As Action Selector
Simple organisms act in accordance to simple stimulus-response pairings, by innate fixed action patterns. More complex organisms, in contrast, actively choose between multiple different response candidates. Such an organism would need a selector which makes this decision. To be effective, a selector must express the following properties
- Singleton. If there are incompatible options, the selector must choose a single behavior. Once a behavior has been activated, the selector must also actively inhibit other behaviors, to promote unanimity of purpose.
- Reward Maximization. The selector must choose the best option, comparing their merits in a common currency.
Redgrave et al (1999) argue that the basal ganglia is a selector, that is has the structural and functional properties that we would expect from such an architecture. The direct pathway disinhibits the winner, the indirect pathway inhibits the losers.
References
- Joel & Weiner (1997). The connections of the primate subthalamic nucleus: indirect pathways and the open-interconnected scheme of basal ganglia-thalamocortical circuitry.
- Nougaret et al (2013). First evidence of a hyperdirect prefrontal pathway in the primate: precise organization for new insights on subthalamic nucleus functions.
- Redgrave et al (1999). The basal ganglia: a vertebrate solution to the selection problem?
