
Part Of: Anthropogeny sequence
Content Summary: 1300 words, 13 min read.
The Domestication Syndrome
Since our emigration out of Africa 70,000 years ago, Homo Sapiens have domesticated many other species, including
- dogs (18 kya, first domesticated in Germany)
- goats, sheep (11 kya)
- cattle, pigs, cats (10 kya)
- llamas, horses, donkeys, camels, chickens, turkeys (5 kya)
- foxes (50 years ago)
Consider the domestication of wolves into dogs. An important part of the environment of a species is other species- not merely its predators or pathogens but its symbionts. In this case, canines began to get food from human campsites. Dogs that were less aggressive were (by unconscious preference and conscious intent) more successful at extracting resources. This process is known as artificial selection.
Most ancient dogs kept by hunter-gatherers share a common body shape. More recently however, humans have conducted pedigree breeding: influencing the morphologies of different dog breeds. We have used this power to sculpt breeds as diverse as the Chihuahua and the Great Dane.
The defining feature of domestication is docility: a reduction in reactive aggression. All domesticated species exhibit this feature, in comparison to their wild counterparts. Not all species are capable of this sort of control. For example, humanity has tried for centuries to domesticate big fauna such as zebras, lions, and hippos. However, some breeds have reproductive and aggressive styles that prohibit domestication.
But domestication doesn’t just bring about a change in behavior. It also brings with it a bewildering number of anatomical changes, to essentially all domesticated species. The domestication syndrome include:
- Docility (agreeableness, reduction in irritability)
- Depigmentation (especially white patches, brown regions)
- Floppy ears
- Shorter ears
- Shorter jaws
- Smaller teeth
- Smaller brains (10-15% reduction in volume)
- More neotenous behavior (juvenile behavior that extends into adulthood).
- Curly tails
Most domesticated species express some aspect of the domestication syndrome, as we can see in the following table:

Three Theories of Domestication
The sheer complexity of the domestication syndrome requires an explanation. What is the link between floppy ears and docility?
Three hypotheses suggest themselves:
- Multiselection. Are the symptoms of domestication all expressions of human preferences? Do we simply like curly tails and floppy ears?
- Environment. Is there something about proximity to humans that incentivizes these changes?
- Byproduct. When the genes for aggression are altered, does that somehow incidentally cause these other changes?
Animal husbandry practices are lost to the sands of time. Nevertheless, there is a way to test multiselection directly: by creating a domesticated species in the laboratory.
In 1959, Dmitri Belyaev began trying to domesticate silver foxes. He used exactly one criterion for selection: he only bred pups that exhibited the least aggression. Skeptics thought it would take centuries to complete the domestication process. But changes in temperament were seen after only four generations. At twelve generations, “elite” foxes began to emerge with dog-like characteristics: wagging their tails, allowing themselves to be petted etc. At twenty generations, the entire population was considered fully domesticated.
Despite only selecting for docility, Belyaev’s foxes exhibited the full domestication syndrome. The foxes inexplicably developed floppy ears, curly tails, white patches, etc etc. The multiselection hypothesis is false.
Is there something about proximity to humans that selects for the domestication syndrome? The environment hypothesis seems false for two reasons.
- First, when they return to the wild, domesticated species take a long time reverting their characteristics. In fact, often domestication gives them a selective advantage over their wild cousins.
- Second, as we will see in the next section, self-domesticated species such as bonobos exhibit the syndrome despite their evolution not being influence by hominids.
The byproduct hypothesis is our only remaining explanation for the domestication syndrome. But what specific system produces these changes?
The Biological Basis of Domestication
In order to fully explain aggression reduction, we must understand it at a biological level.
The primary basis of aggression reduction is a shrinking amygdala and periaqueductal gray (PAG). These modules comprise the negative valence system which learn which stimuli are negatively-valenced, and forward them to the mobilization system (e.g., snake → bad → run away). Serotonin inhibits the negative valence system, and domesticated animals have much high concentrations of serotonin receptors in these regions. Finally, it appears that these changes mostly act across development. The negative valence system comes online only slowly: there exists a socialization window in the first month of a wolf’s life, where it can learn “humans are okay”. Domestication primarily acts by increasing the socialization window from one to twelve months. If a dog isn’t exposed to a human in its first year, it’s now-active fear system will kick in: it will be wild for the rest of its life.
So what biological system is able to a) expand the socialization window, and b) induce the rest of domestication syndrome? The leading hypothesis involves a feature of development called the neural crest.
A blastocyst has no brain. To correct this unfortunate situation, every vertebrate genome contains instruction for constructing a neural tube. This structure emerges via folding.
The neural crest resides between the epidermis and the neural tube. These neural crest cells (NCCs) then proceed to migrate to a certain number of other anatomical structures to assist development. When the NCC migration malfunctions, the resultant disease is called a neurocristopathy. Many neurocristopathies result in outcomes similar to the domestication symdrome! For example, here is the effect of piebaldism:
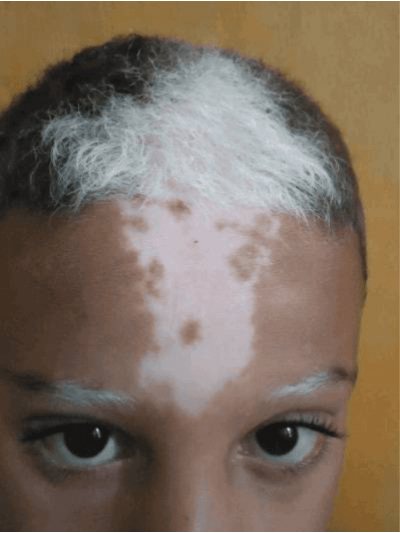
The mild neurocristopathy hypothesis (Wilkins et al, 2014) holds that domestication syndrome is a byproduct of changes to the NCC migration pattern.

The hypothesis, however, is not very detailed (how exactly is NCC migration changed? What are the genomic and epigenomic contributions?). It is more of a promissory note than a mechanistic account. And there are other holistic hypotheses on offer, including genetic regulatory networks (Trut et al 2004) and action of the thyroid gland (Crockford 2000). It seems clear that, in the coming decades, a detailed mechanistic theory of domestication will emerge to vindicate the byproduct hypothesis.
Two Kinds of Domestication
Natural selection explains why the “design requires a designer” trope is obsolete. For the same reason, domestication can occur in the absence of a domesticator. More precisely, change in a species ecological niche can itself select against aggression. Because aggression is very relevant to survival, we see plenty of species that have increased, and plenty that have decreased their rates of aggression. We call those less aggressive species self-domesticated: they became more peaceful in the absence of humans. What’s more, these species also exhibit the domestication syndrome.
Another example is embedded in Foster’s Rule. Islands tend to be geologically more recent than continents, so their populations derive from the continent rather than vice versa. Islands tend to have fewer predators, but also fewer resources. Reduced predation increases the size of small animals (e.g., dodos evolved from pigeons), but limited resources decreases the size of big animals (e.g. the 3ft tall dwarf elephant).
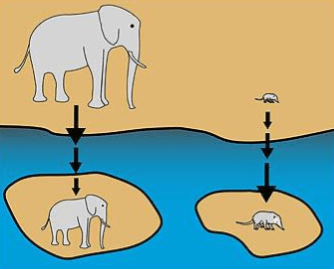
Because islands have fewer predators, they also tend to have higher population densities; as such, reactive aggression is a less useful strategy. Selection favors the less aggressive. And we can see the domestication syndrome in island species. For example, the Zanzibar red colobus monkey has diverged from the continental red colobus along the same trajectory as dogs diverged from wolves.
Other examples of self-domestication can be found with group size reduction (ungulates, seals) and low-energy habitats (extremophile fish).
Finally, bonobos provide a particularly relevant example of self-domestication. Because food is more plentiful (don’t have to compete with gorillas for vegetation), females can spend time close to one another. Proximity produces bonding, and female coalitions exert pressure on bonobo behavior.
- In chimps, bullying women increases reproductive success. Chimps will systematically beat up all females in their group as a coming-of-age ritual.
- In bonobos, female coalitions retaliate against male aggression, making it unprofitable. Sexual selection then acts against reactive aggression.
So we can see that domestication (i.e., reduction in aggression) can come in two flavors: traditional vs self-domestication.
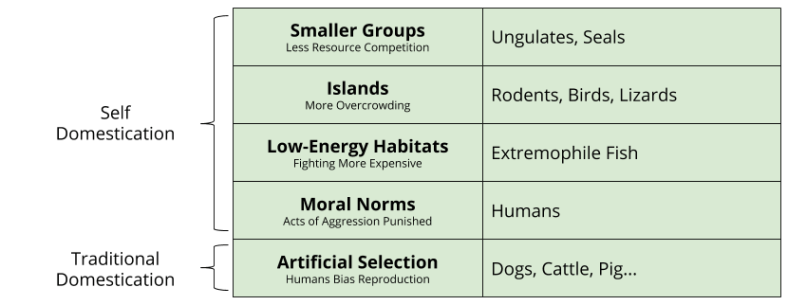
As we will see next time, Homo Sapiens is yet another example of a self-domesticated species. See you then!
Related Resources
- Wilkins et al (2014). The “domestication syndrome” in mammals: a unified explanation based on neural crest cell behavior and genetics

Cool – I hadn’t heard of the domestication selection theory relating to neural crest development- it will be interesting to see whether it holds up as more investigations take place. The traits of domestication do seem a strange collection fur/skin patterning / floppy ears / curly tails – seen across diverse species (e.g. dogs / pigs / rabbits). It does suggest a common cause that is selected for by either humans or the environment.
LikeLike
Very much agree! My betting money is that it will be replaced with a far more specific mechanism (but plausibly still implicating neural crest subprocess).
IOU (and my other readers) a post of self-domestication of sapiens. Until I do, recommend Wrangham *Goodness Paradox*
LikeLike
“In chimps, bullying women increases reproductive success. Chimps will systematically beat up all females in their group as a coming-of-age ritual.”
Bullying women increases reproductive success of both male and female chimps?
LikeLike
Sorry for the belated reply, was struggling to find the reference. Here’s Stanford (2018) The New Chimpanzee pp43.
> Dominance in chimpanzees is not about physical size. Each sex has a dominance hierarchy, and all adult males are usually dominant to all females. Adolescent males do not typically exhibit the hallmarks of a dominance struggle (Sandel et al 2016). They don’t submissively pant grunt to one another, and dominance among them is hard to discern. But once a young male reaches adulthood, he begins his climb to higher rank by taking on and intimidating each adult female. When he has risen in status above the most dominant female, the young male finds himself at the bottom of the male dominance network.
Female dominance doesn’t have this feature.
LikeLike
Thank you! It’s a curious passage to me. I’m a bit surprised by the wording, which seems to imply that this behavior is basically invariant across chimpanzee societies. Makes me wonder whether it really is, and why certain patterns might occur over and over again, or when they might break, or when there might be a larger range of behavior, in groups of animals.
LikeLiked by 1 person
I’m currently exploring the topic of dominance hierarchies; found a couple other relevant resources you may enjoy, De Waal (1982) Chimpanzee Politics paints a complementary picture, following the coming of age of adolescent Nikkie. This is from Boehm (1999) Hierarchy in the Forest:
> Every young male, as he approaches or reaches adolescence, becomes driven by political aspirations. First, he displays at low-ranking adult females until they begin to pant-brunt submissively when they greet him. Then he moves on to the more formidable females. Sometimes he suffers reverses along the way, particularly if the females have allies to help them, Eventually he will dominate all the females and begin to direct his displays at lower-ranking adult males. (Goodall 1986, 1990). If he is successful in that pursuit, he keeps working his way up the male hierarchy until he can go no further. At Gombe Mustard and Jomeo never made it very far, but even they were dominant to all the adult females.
Of course, patterns of dominance and gender vary quite widely across species. Since hominins diverged from chimps towards a) multilevel societies and b) cooperative breeding, it would be interesting to know how dominance structures & gender relations respond under those social conditions.
LikeLike