
Part Of: Anthropogeny sequence
Followup To: The Walking Ape
Content Summary: 2100 words, 21 min read
A Brief Review of Human Evolution
The most recent common ancestor of humans and chimpanzees lived 7 mya (million years ago). The very first unique hominin feature to evolve was bipedality, which was an adaptation for squat-feeding. The australopiths were bipedal apes. They could walk comfortably, but retained their adaptations for tree living as well. Dental morphology and microwear together suggest that australopiths acquired food from a new source: tubers (the underground storage organs of plants).
Climate change is responsible for the demise of the australopiths. Africa began drying out about 3 million years ago, making the woodlands a harsher and less productive place to live. Desertification would have reduced the wetlands where australopiths found fruits, seeds, and underwater roots. The descendents of Australopithecus had to adapt their diet.
The paranthropes adapted by promoting tubers from backup to primary food. These impressive creatures comprise a blend of human and cow-like features. In contrast, the habilines (e.g., Homo Habilis) took a different strategy: meat eating. These creatures had the same small bodies, but larger brains. Their hands show adaptations for flexibility, and their shoulders and elbows for throwing missiles. They began making stone tools (Mode 1 tools, the Oldowan industry). They presumably used these anatomical and cultural gifts to compete with other scavengers on the savannah (projectiles to repulse competitors, stone flakes to speedily butcher a carcass).
The habilines in turn gave rise to
- [1.9 mya] The erects (H erectus) with near-modern anatomies.
- [0.9 mya] The archaics (H heidelbergensis) appear, who eventually give rise to the Neanderthals, Denisovans, and us.
- [0.3 mya] The moderns (H sapiens) emerge out of Africa, and completely conquer the globe.
A Closer Look
Yes, humans are apes. But why do we look so different from our closest living relative, the chimpanzee?
I have previously explained why we are bipedal (flexible waist, straight backs, walking on two feet).
But why do we have scent glands in our armpits? Fat in our asses? Such weird hair? Hairless skin with massive subcutaneous fat deposits?
Most of these changes were introduced with Homo Erectus:
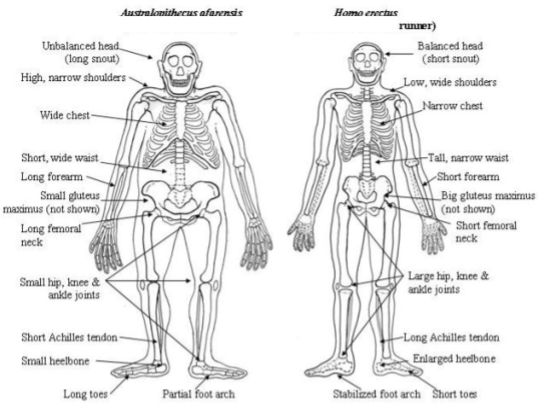
Natural selection explains why bodies change. Anatomical innovations are selected when they enable more efficient exploitation of some particular niche.
So what ecological niche forged the modern human body?
Where Homo Erectus Evolved
The australopiths never made it beyond the southern margins of the Sahara. Because the adaptation of equatorial species inhibits their colonization of temperate regions, the successful emigration of the erects out of Africa strongly suggests that this was a northern, not a tropical species.
To evolve adaptations to dry, open country, the erects would have had to suffer a period of isolation from other hominins, in an appropriately discrete habitat. There were few, perhaps no, places in tropical or Southern Africa that could have provided such a combination. Comparing these constraints with the distribution of Homo Erectus fossils, comparative zoologist Jonathan Kingdon submits there the two most plausible contenders where the erects could have evolved are the Atlas Mountains, or Arabia.
Nasal evidence corroborates the hypothesis that they evolved in a desert environment. The entry to the primate nasal passage is flat, with straightforward air intake. Erect skulls show the first evidence of a protruding nose. A protruding nose forces the air at a “right angle” before entering the nasal cavity.
One of the responsibility of the nasal passage is to humidify the air before it is passed to the lungs. The increase in room and turbulence serves to amplify the humidification of inhaled air. Our noses are adaptations for desert living (Pontzer et al 2021).
A New Thermoregulation System
There are two things unique to human skin:
- Functional hairlessness. We modern humans have hair, but it is so thin compared to chimpanzees that we are effectively hairless.
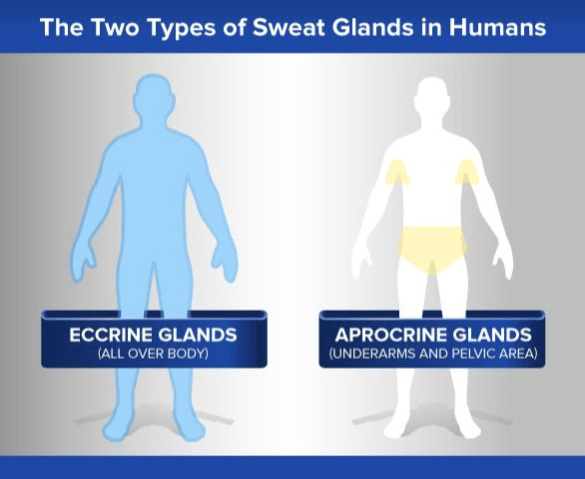
- Eccrine sweat glands. Our skin also contains a novel approach to sweat glands.
These two features are linked: we now know in exquisite molecular detail how incipient hair follicles are converted into eccrine glands (Lu et al 2016).
Other primates rely on oil-based apocrine sweat glands. The emergence of water-based eccrine glands in humans led to the “retirement” of apocrine glands in our lineage. The distribution of odor-producing apocrine glands was ultimately confined to our underarms and pubic regions.
Losing our hair had two important side-effects:
- Skin pigmentation. Fur protects against ultraviolet radiation. Without it, melanin was used as an alternate form of natural sunscreen.
- Why do otherwise-bald humans have hair at the tops of their heads? This is the location of maximal radiation.
- Why didn’t all humans remain dark-skinned? Melanin also inhibits the skin’s production of Vitamin D, and different locales have different radiation levels, requiring new tradeoffs to be struck.
- Subcutaneous fat. Ever seen a hairless chimpanzee? Human skin is much less wrinkled than other skin. Why? Even in non-obese people, humans store more of their body fat below the skin (versus in the abdomen, or between the muscles). This change has three complementary causes:
- carnivores tend to store fat in this way,
- mitigate the hernia risk associated with bipedality
- replace the insulation services of fur, without interfering with sweat system.
We have reviewed four changes in human skin. Rather than a discrete event, these changes presumably evolved gradually, and in tandem.

Yes, but why are we hairless? There are many competing theories.
Jonathan Kingdon claims these skin adaptations arose late, as a parasite avoidance mechanism induced by increased population densities. Two rationales are provided: hair is a potent vector of infection, and the eccrine sweat system also has antibiotic properties.
This interpretation is challenged by genetic evidence that shows hominins were naked at least 1.2 mya, if not earlier (Rogers et al, 2004).
However, given the evidence suggesting Homo Erectus evolved in a desert climate, the most parsimonious theory seems to involve thermoregulation. We were exposed to less direct radiation given our upright posture; fur no longer served as critical of a role. But the overall climate was warm and dry,
Humans as Cursorial Species
A cursorial animal is one that is adapted for long-distance running, rather than animals with high acceleration over short distances; thus, a leopard is considered cursorial, while a cheetah is not. Other examples include wolves, horses, and ostriches.
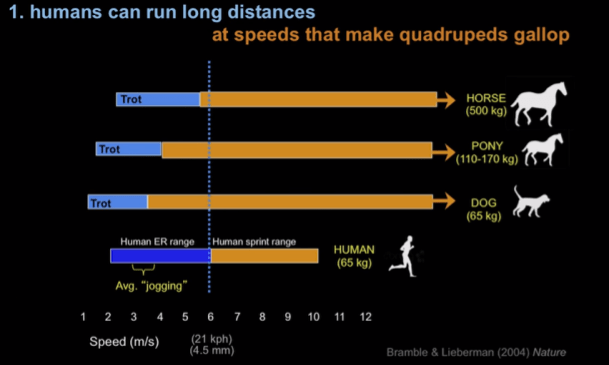
Fit human amateurs can regularly run 10 kilometers, and longer distances such as marathons (42 kilometers) are achieved by tens of thousands of people each year. Such distances are unknown if not impossible for any other primate, but are comparable to those observed in specialized mammalian cursors on open habitats. African hunting dogs, for example, travel an average 10km per day.
Racing horses can gallop 10 kilometers at 9 meters per second. However, the sustainable galloping speeds in horses decline considerably for runs longer than 10-15 minutes. Well-conditioned human runners exceed the predicted preferred galloping speed for a 65-kg quadruped, and can even occasionally outrun horses over extremely long distances.
Thus, despite our embarrassingly slow sprinting speed, human beings can outcompete even cursorial animals at endurance running over large distances. How come? The answer has to do with our unique cooling system.
When other mammals trot, they cool themselves by panting. However, above certain speeds a quadruped transitions to a full gallop, which precludes panting. A horse can trot all day, but it cannot gallop continuously without overheating.
Human adaptations for running, and our unique eccrine sweat-based cooling system, meant that humans have a larger trot/gallop (jog/sprint) transition threshold. Our superior cooling technology is accentuated in high heat. We are literally the only mammal that can run a marathon in high heat.
Why are we Born to Run?
Our bodies are designed for endurance running. We are cursorial animals. But why?
To achieve this, hominids exploited a new form of predation called persistence hunting. The most successful persistence hunts will involve:
- Time: middle of the day (during peak heat)
- Target: big prey (overheats faster)
If you chase a big animal above its trot/gallop transition speed, the animal will easily distance itself and begin panting. But you can track the animal, and chase it again before it has the opportunity to fully recover. Repeat this process, and after 10-25 km you will successfully drive the prey into hyperthermia. This style of hunting has a remarkable 75% success rate. Modern hunters typically prefer to use the bow and arrow, but persistence hunting is still in their repertoire. Before the invention of projectile weapons some 71 kya, persistence hunting surely played a larger role.
We know that habilines ate meat (many bones show signs of their butchery). But they likely acquired meat by scavenging, as they were not particularly effective carnivores. Their adaptations for projectiles were presumably used to repulse competitors, and stone tools certainly helped speedily butcher a carcass.
Of the dozens of running adaptations in our Homo Erectus, a substantial fraction already exist in habilines. Presumably the re-invention of our skin had begun too. These processes presumably began for simple reasons (it pays to move quickly, and have less fur, in the savannahs that emerged 3 mya).
Persistence hunting completely changed the game. Adaptations for running brought steep rewards. In a typical persistence hunt, the hunter averages an energy expenditure of 850 Kcal; they energy gains from big game is multiple times larger. Compare the calorie budget for a modern-day hunter-gatherer with that of chimps: in our prime, we produce twice as many calories as we consume!
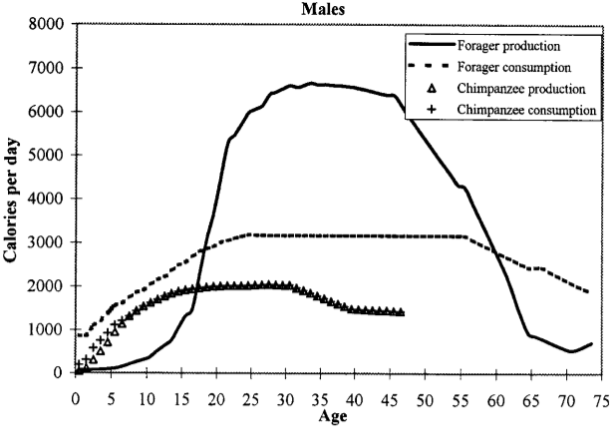
Life is fundamentally about getting energy to make more life.
Persistence hunting was the turning-point in human evolution. Our species began winning, in terms of our reliably acquiring surplus energy. This surplus was the reason why our lineage could “afford” bigger brains, taller bodies, more frequent births, and longer childhood. All of these characteristics have improved gradually & continuously since the erects emerged.
Our Cursorial Adaptations
We have looked at the reasons behind our running. What does anatomy tell us?
First, let’s compare the physics of walking vs running:
- Walking is an inverted pendulum mechanism. Our feet and our hips alternate as the center of rotation.
- Running is a mass-spring mechanism. Ligaments transfer foot-strike kinetic energy into tendons, which is released as we bounce onward.
Walking doesn’t require springs – but running does. And the bodies of erects have two new ligaments that serve precisely this purpose:
- The Achilles’ tendon stores and releases 35% of energy expended while running (but not walking). In chimps, this tendon is 1cm long. In erects, it is 10cm and much thicker.
- The dome-shaped arch of the foot is another spring, which lowers the cost of running by 17%.
During bipedal running the risk of falling and sprained ankles is high, which in the ancestral environment had adaptive consequences. Thus, the human body also developed many stabilization techniques:
- Gluteus maximus. Barely active during walking, this muscle contracts forcefully during running to prevent the trunk from toppling forward.
- Various head stabilization devices. Promotes vision continuity and protects the brain (watch a runner with a ponytail sometime).
- Enlarged semicircular canals (balance organs) in inner ear, which can be seen by measuring certain dimensions of fossilized skulls.
I have listed five features of our anatomy that relate to endurance running. Lieberman et al (2006) list twenty:

As you can see, not all of these running adaptations emerged with Homo Erectus. Homo Habilis already shows adaptations for running. It would not surprise me in the slightest if that species also saw the beginnings of our skin trajectory.
Adaptations for running came at a price. We have lost our ability to climb trees. We are the first primate to lose this ability.
Takeaways
Why do humans look so different from our closest living relative, the chimpanzee?
Why do we have scent glands in our armpits? Fat in our asses? Such weird hair? Hairless skin with massive subcutaneous fat deposits?
Animal body plans are designed to excel in a particular niche. Our bodies are designed for persistence hunting. Compared to other primates, our anatomies optimize for thermoregulation, efficient energy transfer, and stabilization during running.

Chimpanzees don’t need to exercise to stay fit. We do. Our health sees dramatic benefits from aerobic exercise, especially running.
References
- Bramble & Lieberman (2004). Endurance running and the evolution of Homo
- Lieberman et al (2006). The human gluteus maximus and its role in running
- Lu et al (2016). Spatiotemporal antagonism in mesenchymal-epithelial signaling in sweat versus hair fate decision.
- Pontzer et al (2021). Evolution of water conservation in humans
- Rogers et al (2004). Genetic Variation at the MCiR Locus and the Time since Loss of Human Body Hair

{kind=link}
Great job.
One more step to take, in your analysis, imho….
Invert the question of the hairless ape: when not running and cooling, how did the naked ape offset the cooling efficiencies? Desert environments aren’t just hot; they’re also frequently cold. This is where be wrangham and kingdom meet. Also, clothing.
It’s useful to tie it all together, I think.
LikeLike
I actually haven’t quite understood your comment yet! Could you elaborate?
I mentioned subcutaneous fat (here) and fire (another post) as important tools erects used to thermoregulate. Did you have something else in mind?
(Sewn clothing originates 70 kya, Fur scraping and body louse evidence go back to between 500 to 170 kya. Evidence seems to suggest we were naked & unclothed for a long stretch of deep time)
LikeLike
That consumption\production graph is something I don’t recall seeing before. It strongly suggests a need for social cooperation where the excess from prime foragers is required to fulfill the caloric needs of others. Interesting to see how the evolution of all these different traits interact.
LikeLiked by 1 person
Yes, absolutely. Technical keyword for this phenomenon is provisioning. I’m hoping to someday explore that topic. In the meantime,
Provisioning becomes even more remarkable when you consider female calorie budget in *A Theory of Human Life History Evolution: Diet, Intelligence, and Longevity*
Wrangham In *The Cooked and the Stolen* suggests that cooking was the asymmetry that provoked provisioning & sexual division of labor (male hunter female gatherer). But Kuhn and Stiner *What’s a Mother to Do?
The Division of Labor among Neandertals and Modern Humans in Eurasia* suggest that while Neanderthals had provisioning and pair-bonding, they show zero evidence for division of labor.
LikeLiked by 1 person