
Part Of: Neuroanatomy sequence
Content Summary: 2200 words, 22 min read
Four Cortical Networks
Cognitive neuroscience typically employs fMRI scans under a carefully crafted task structure. Such research localized various task functions to different neural structures (cortical areas). For example, these studies produced evidence suggesting that the hippocampus is the seat of autobiographical memory.
In the early 2000s that researchers stumbled upon a different question, what brain regions are active when the brain is at rest? Here is Raichle (2015) describing his discovery of the default mode network.
One of the guiding principles of cognitive psychology at that time was that a control state must explicitly contain all the elements of the associated task other than the one element of interest (e.g., seeing a word versus reading the same word). Using a control state of rest would clearly seem to violate that principle. Despite our commitment to the strategies of cognitive psychology in our experiments, we routinely obtained resting-state scans in all our experiments, a habit largely carried over from experiments involving simple sensory stimuli, in which the control state was simple the absence of the stimulus. At some point in our work, and I do not recall the motivation, I began to look at the resting-state scans minus the task scans. What immediately caught my attention was the fact that regardless of the task under investigation, the activity decreases almost always included the posterior cingulate and the adjacent precuneus.
Well before the discovery of the default mode network, Peterson and Posner (1980) had put forward three networks underlying attention. The dorsal attention network generated salience maps across the perceptual field, and used these maps to orient to interesting stimuli. The ventral attention network is involved in attention switching to novel stimuli. The executive network produces top-down control of attention, for example translating the instruction “pay attention to the green triangle” to sustained attention on an otherwise-uninteresting object.
Fox et al (2005) brought these two worlds together in their seminal paper, which identified a brain-wide task-positive network which anti-correlated with their task-negative network. Their use of resting-state functional connectivity MRI (rs-fcMRI) provided independent evidence of the existence of these networks. By examining the cross-correlations of “noise” in the BOLD signal, one can identify regions that “fire together”, and may be functionally integrated.

Fox et al’s task-negative network was the default mode network. And the task-positive network seemed to contain two networks previously identified: the executive network, and the dorsal top-down attention network. The ventral attention network, however, was not identified in their analysis.
And that was the state of the world in 2006. Neuroscientists had identified four networks, which we will henceforth call intrinsic connectivity networks (ICNs). They are:
- Executive Control
- Dorsal Attention
- Ventral Attention
- Default Mode Network
Towards Eight Networks
While the data supporting the legitimacy of these networks was strong, these anatomical structures pose a fairly routine challenge in neuroscience: they correlate with “too many functions”. Take the default mode network. It is associated with mind-wandering, social cognition, self-reference, semantic concepts, and autobiographical memory. How could one structure produce these widely divergent behaviors?
In the case when you have too many functions, you have two options: look for more specific mechanisms (Q3), and group similar concepts (Q4). In many neuroscience applications, the former is more productive: reality has a surprising amount of detail.

Researchers began to find subnetworks within the executive control.
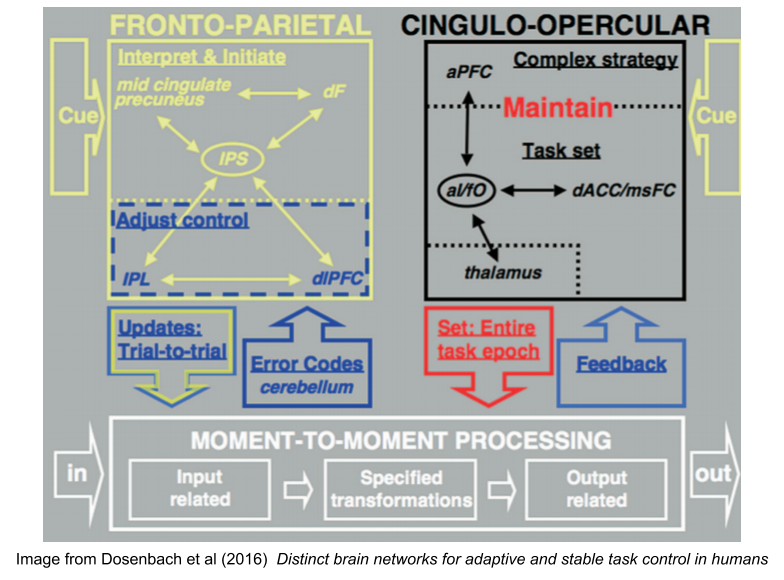
Dosenbach et al (2007) found two networks within the “executive network”. They found a fronto-parietal control network (FPCN), involved in error correction, and control over task execution. They also found a cingulo-opercular control network (COCN), involved in task set maintenance. The FPCN was most active at task onset and errors, the COCN expressed activity consistently throughout the task.
These subgraphs usefully pick out useful psychological concepts. We have long known that rehearsal increases working memory capacity from 3 to 7 chunks. It seems the COCN produces this miracle (but recall that the contents of working memory, the stuff it rehearses, lives in perceptual cortex, Postle 2006). Likewise, psychologists have long studied the phenomenon of willpower or volition. The FPCN might be the neural substrate of this ability.
Seeley et al (2007) also found substructures within the original executive network. But they didn’t see a rehearsal system in the cingulo-opercular regions. Instead, they found a salience network, which bound affective and emotional information into perceptual objects, and links to the basal ganglia reward system.
Since publication, each of these networks have been replicated dozens of times, using a widely diverging set of paradigms (ROI vs voxel granularity, fMRI vs rs-fcMRI) and statistical techniques (graph theory, dynamical causal modeling, hierarchical clustering, and independent component analysis).
Unfortunately, these subnetworks looked and behaved radically differently. For years, neuroscientists collected data using these diverging theories. Peterson & Posner (2012) updated theory of attention rely on Dosenbach’s rehearsal network, whereas many other articles took inspiration from Seeley’s salience network.
And then, a miracle. Power et al (2011), using graph theoretic tools and more granular data, identified both salience and rehearsal networks hidden within the cingulo-opercular graph. Despite the close proximity of these two networks, they perform dramatically diverging functions (left image).
They also discussed the spatial distribution of these networks across cortex. Essentially, the attention networks are sandwiched between sensorimotor networks and prefrontal control networks. This configuration might play an important role in reducing wiring cost for between-network communication.

This work was largely replicated in Yeo et al (2011), which in contrast to the biological tack of Powers et al (2011), used a more statistically-oriented approach.
ICNs are not exclusive to cortex. Habas et al (2009) found strong links between cerebellar substructures and various ICNs. Ji et al (2009) find correspondences between other midbrain structures (e.g., various nuclei in the amygdala).
Default Mode Network and Interoception
Power et al (2011) also compared network properties of their ICNs and discovered two categories of ICN:
- processing networks that are directly involved in perceptual-action loops. These networks tend to be very modular in their organization.
- control networks that modulate cybernetic loops. These networks tend to have more extra-subgraph relationships.

The above illustrates an intriguing finding: the default mode network is a processing network, rather than a control network. But what sense modality does underlie?
The answer is straightforward to an affective neuroscientist. The default mode network and the salience network comprise the seat of the hot loop; it performs:
- interoception (viscerosensory body perception); and
- allostasis (visceromotor body regulation)
It is a cornerstone of dual cybernetic loops. Indeed, comparative studies with macaque monkeys put empirical meat on this assertion:
- { anterior cingulate cortex, dorsal amygdala, ventral anterior insula } perform visceromotor functions (allostasis)
- { dorsal anterior insula } perform viscerosensory functions (interoception).
As Kleckner et al (2017) show, these assertions are born out by myriad human rs-fcMRI studies, and further bolstered by tract-tracing studies in non-human animals.
I’ll note in passing that most experts now detect three subgraphs within the default mode network (cf Andrews-Hanna et al 2014). But the functional signature of these subgraphs has not yet been worked out, so let me simply note this development in passing.
Network Neuroscience
We have so far discussed results from function-derived structures, with techniques such as rs-fcMRI computing ICNs from the dynamics of neural activity. A complementary research tradition can be described as anatomy-derived structures, which is a more anatomical emphasis on connectome studies. These two network types have important differences, including time scales (anatomy-derived structures tend to persist longer than task-dependent structures) and levels of detail (neuron versus region-of-interest). Nevertheless, these data can be made to usefully constrain one another (functional networks are beginning to look more like structural networks, and vice versa).

These approaches have recently coalesced (Basset & Sporns 2017) into the new discipline of network neuroscience. Very similar techniques are used in network science and social network analysis in the analysis of social networks.
If a neuron is a node in a graph, and a synapse is an edge, what properties does the graph of a human brain enjoy? There are several kinds of networks possible. Regular networks enjoy rich local connections, but few cross-graph connections. Random networks enjoy more long-term connections, but are less structured. Small-world networks represent a kind of middle ground, with lots of local structure but also afford the ability to make long-term connections.
With graph theoretic measures, we can quantitatively partition networks into sets of modules. A hub is a node with high degrees of centrality (e.g. node degree: how many edges that node supports). A connector hub facilitates between module communication; a provincial hub promotes communication within modules.

Connectome studies (anatomy-derived structural networks) have shown that brain hub regions are more densely interconnected than predicted on the basis of their degree alone. This set of unusually central connector hubs is called the rich club. The rich club is the most metabolically expensive areas of cortex: they are “high cost, high value”. They are loosely analogous to DNS servers (the thirteen servers are the global basis of the internet)
Human neural architecture is thus a specific kind of small-world network, one equipped with a “rich club”. These topologies have been shown to exist in other species, such as macaque monkeys and cats. Interestingly, some hubs (posterior cingulate, precuneus, and medial frontal cortex) act as sinks (more afferent than efferent connections) whereas and hubs within attentional networks (incl. dorsal prefrontal, posterior parietal, visual, and insular cortex) act as sources (more efferent than afferent connections).
What does this have to do with ICNs? As shown by von den Heuval & Sporns (2013b), the rich club seems to be the substrate of inter-ICN communication.

Networks vs Consciousness
According to global workspace theory, consciousness contents are generated via a publicity organ which selects perceptual information worthy of further processing by downstream modules. There is, however, much disagreement about the mechanism of conscious contents. Theories include:
- Dehaene and Changeux have focused on frontal cortex
- Edelman and Tononi on complexity in re-entrant thalamocortical dynamics
- Singer and colleagues on gamma synchrony
- Flohr on NMDA synapses
- Llinas on a thalamic hub
- Newman and Baars on thalamocortical distribution from sensory cortex
Shanahan (2012) offered a new hypothesis, that the rich club has recently hypothesized as the basis of consciousness. Its central location and role synchronizing large-scale brain networks makes it a plausible suspect. However, it is unclear whether the rich club is primarily facilitated by corticocortical white matter, or corticothalamic reentrant loops. If the latter, the hypothesis would converge with existing theories that emphasize the role of the thalamus.
Traits and Individual Differences
Researchers are becoming increasingly aware of individual differences across brain regions. Here, for example, is the group-average ICNs contrasted with a single individual:

Seitzman et al (2019) describe clusters within these individual differences.
- Most network variance is at the individual level (less so between tasks, or over time).
- Most individual ICNs fall into two distinct trait-like variants: one with large DMN, the other with large FPN.
- It was the network assignment of variants, rather than their anatomical location, that differentiate the variants.
- ICN variants occur most often in associative networks, and most often in the right hemisphere.

At this time, it is hard to say what behavioral or personality differences are driven by these variants.
Networks vs Modules
ICNs comprise a central organizing principle of the nervous system. But they are not the only such principle; we have identified some fifteen others!

It is difficult to reconcile intrinsic connectivity networks (ICNs) with massive modularity, so that will be the topic of this section.
ICNs have been seized upon by some theorists in the Bayesian predictive coding traditions (e.g. Barrett & Simmons 2015) as evidence of the illegitimacy of modules. But most ICN theorists still admit the centrality of modules (e.g., Sporns & Betzel 2015). Here, for example, is von den Heuval & Sporns (2013a):
Since the beginning of modern neuroscience, the brain has generally been viewed as an anatomically differentiated organ whose many parts and regions are associated with the expression of specific mental faculties, behavioral traits, or cognitive operations. The idea that individual brain regions are functionally specialized and make specific contributions to mind is supported by a wealth of evidence from both anatomical and physiological studies. These studies have documented highly specific cellular and circuit properties, finely tuned neural responses, and highly differentiated regional activation profiles across the human brain. Functional specialization has become one of the enduring theoretical foundations of cognitive neuroscience.
Most researchers now admit the interaction of both principles (specialization and integration). It is unclear how it could be otherwise. I have personally read far too many papers that have described activity in the dorsolateral prefrontal cortex as task-specific, without considering it is a simple expression of the volitional control or working memory rehearsal networks. Similarly, I have read dozens of reviews of the anterior insula that would have profited from the realization that it participates in at least three different ICNs.
The three streams hypothesis integrates notions of massive modularity, cortical streams, the abstraction hierarchy, and the cybernetic loop hypothesis. It is less clear how ICNs might integrate with these organizing principles.
Does the ventral temporal parietal junction (vTPJ) only perform integrative functions in service of the ventral attention network (VAN)? Or does the real estate claimed by these ICNs also used to perform specialized computations such as mindreading? The latter proposition strikes me as more likely. But I’d like to see more data on this. To be continued…
Wrapping Up
The human cortex has intrinsic connectivity networks (ICNs) that coordinate to provide integrative services on behalf of our central nervous system. Researchers have so far identified the following networks:
- Default mode network (DMN) and its three subnetworks described in Andrews-Hanna et al 2014
- Ventral Attention Network (VAN), reinterpreted by Ji et al 2019 as the language network.
- Dorsal Attention Network (DAN)
- Fronto-Parietal Control Network (FPCN) implicated in volitional control, and per Duncan et al 2020, possibly fluid intelligence.
- Cingulo-Opercular Control Network (COCN)
- Salience Network, reinterpreted by Ji et al 2019 as a subnetwork within COCN.

A recent reanalysis by Ji et al (2019) adds three more networks into the mix:
- Posterior Multimodal (PMM), possibly involved in spatiotemporal and narrative cognition.
- Ventral Multimodal (VMM), possibly involved in semantic categorization.
- Orbito-Affective (ORA), definitively linked to reward processing.
Until next time.
Works Cited
I’ve put the papers I found especially helpful in bold.
- Andrews-Hanna et al (2014). The default network and self-generated thought: component processes, dynamic control, and clinical relevance.
- Bassett & Sporns (2017). Network Neuroscience
- Barrett & Simmons (2015). Interoceptive predictions in the brain.
- Christoff et al (2016). Mind-wandering as spontaneous thought: a dynamic framework
- Dosenbach et al (2007). Distinct brain networks for adaptive and stable task control in humans
- Duncan et al (2020). Integrated Intelligence from Distributed Brain Activity.
- Fox et al (2005). The human brain is intrinsically organized into dynamic, anticorrelated functional networks
- Gordon et al (2018). Individual-specific features of brain systems identified with resting state functional correlations
- Kleckner et al (2017). Evidence for a large-scale brain system supporting allostasis and interoception in humans.
- Laird et al (2011). Behavioral Interpretations of Intrinsic Connectivity Networks.
- Habas et al (2009). Distinct Cerebellar Contributions to Intrinsic Connectivity Networks
- Ji et al (2019). Mapping the human brain’s cortical-subcortical functional network organization
- Peterson & Posner (1990). The attention system of the human brain
- Peterson & Posner (2012). The Attention System of the Human Brain: 20 Years After
- Postle (2006). Working memory as an emergent property of the mind and brain.
- Power et al (2011). Functional Network Organization of the Human Brain
- Raichle (2015). The Brain’s Default Mode Network
- Seeley et al (2007). Dissociable Intrinsic Connectivity Networks for Salience Processing and Executive Control
- Seitzman et al (2019). Trait-like variants in human functional brain networks
- Shanahan (2012). The Brain’s Connective Core and its Role in Animal Cognition
- Sporns & Betzel (2015). Modular Brain Networks
- Von den Heuval & Sporns (2013a). Network hubs in the human brain
- Von den Heuval & Sporns (2013b) An Anatomical Substrate for Integration among Functional Networks in Human Cortex
- Yeo et al (2011). The organization of the human cerebral cortex estimated by intrinsic functional connectivity
