
Part Of: Sociality sequence
Followup To: Counterfactual Simulation
Content Summary: 1600 words, 16 min read
A Brief Review
Mindreading (also known as mentalizing, the intentional stance, or theory of mind) is the penchant of animals to represent the mental lives of one another. What are the beliefs and desires of those around us? A classic demonstration of mindreading comes from Heider & Simmel (1944):
While the mindreading faculty was designed to understand the minds of other animals, it had no trouble ascribing beliefs and goals to two dimensional shapes. This is roughly analogous to your email provider accepting a tennis ball as a login password.
Another classic demonstration of mindreading is the Sally-Anne test, from Baren-Cohen et al (1985):
Imagine Sally puts a marble in the basket, then leaves the room. Anne moves the marble from the basket into the box. When Sally returns to the room looking for the marble, where will she turn next?
This test is designed to test the ability to understand false beliefs.
The Phylogeny of Mindreading
Historically, mindreading intrigued researchers because it seemed a uniquely human capacity. Here is a quote from Carruthers (2011):
Until 2000, all the evidence seemed to point to the conclusion that apes lack any understanding of the perceptual access of others. But a breakthrough came when it was realized that all the initial experiments had involved cooperative paradigms of one sort or another. For example, the apes might have had to decide which of two humans to request food from. But non-human apes are not naturally cooperative. They are, however, immensely competitive. Hence it might be they weren’t sufficiently motivated to take account of others’; perceptions in cooperative contexts, but would do so in competitive ones.
In a ground-breaking series of experiments, Hare et al (2000, 2001) set out to test this idea.. Indeed, in competing for food, the subordinate seemed to take account of what the dominant could see. Taken together with the results for chimps understanding desires, it seems that monkeys and apes possess at least a goal-perception mindreading system.
Indeed, Call & Tomasello (2008) reviews mindreading studies conducted on chimpanzees, conclusively showing that chimpanzee behavior is responsive to the mental states they perceive in other living things.
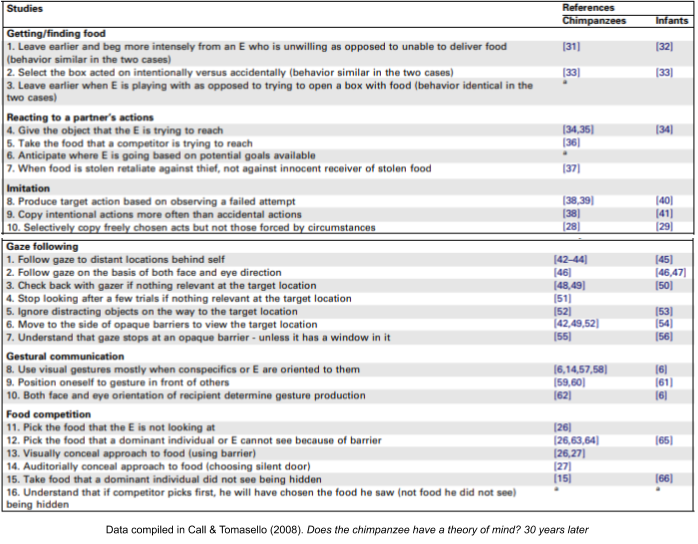
But do chimpanzees possess knowledge of false beliefs? The answer as recently as 2014 was a clear “no”. Dozens of attempts to test chimpanzee awareness of false beliefs had failed to establish any such confidence…
Until they did. Krupenye et al (2016) and Buttelmann et al (2017) demonstrate that, indeed, great apes can pass false belief tasks when the experimental paradigm is sufficiently motivating. In anticipatory looking studies such as this, the question is whether the ape anticipates the action according to what he knows, or what the actor falsely beliefs. As you can see, the ape anticipates the actor to act erroneously (red dots indicate where the chimp is looking)
Chimpanzees (and other great apes) possess a full capacity for mindreading. There is (more contentious) evidence of mindreading in non-ape primates, canids and corvids. I suspect some of the simpler capacities like detecting self-generated motion are deeply homologous. But the more sophisticated faculties seem likely to be cases of convergent evolution. In the case of the great apes, I suspect that their robust mindreading faculties evolved in service of competitive foraging efficiency.
The Ontogeny of Mindreading
Ontogeny builds individuals. It is one thing to say adults can pass these three dozen tests related to mindreading. It is another to say when the ability to pass these tests is delivered by ontogeny.
For example, when are children able to pass the Sally-Anne test? Baren-Cohen et al (1985) demonstrate that this ability arrives at 44 months, except in autistic children for whom the ability to pass is severely delayed.
Do the Sally-Anne results suggest that the ability to model the knowledge of others arrives at 44 months? By no means! While a child can verbalize false beliefs at 44 months, it turns out their subconscious mind begins generating the appropriate expectancies much earlier. Looking time studies (e.g., Onishi & Baillargeon 2006) demonstrate that 15 month infants are surprised by violations of false-belief scenarios.
Why the 2.5 year gap between intuition-based and language-based understanding of false belief? Carruthers (2011, section 8.4) reviews attempts to explain it; but I didn’t find his explanation (or anyone else’s) particularly satisfying. It turns out that this intuition vs report gap may generalize beyond mindreading; similar phenomenons occur in e.g., measures of outcome bias. Let’s bookmark this as an open question, and move on…
While the mindreading abilities of adult humans is fairly comparable to other apes, in humans it is more frequently expressed in non-competitive contexts. Further, comparative development data has shown that mindreading emerges much earlier in human infancy:
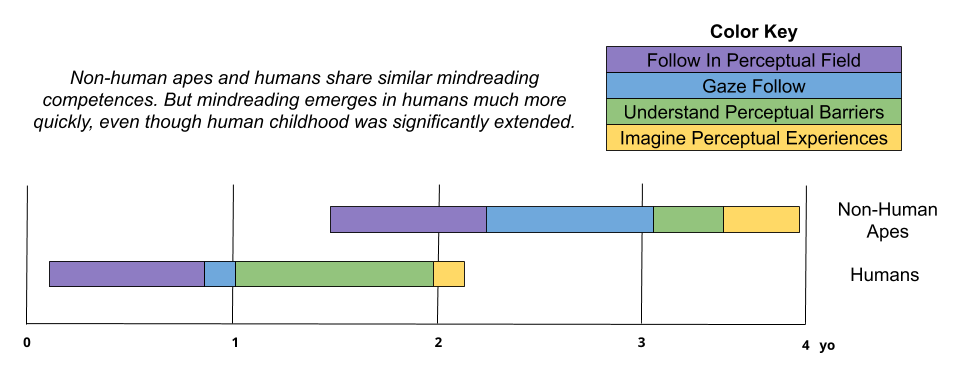
As we will see later, this acceleration of mindreading has important implications.
Sub-processes: Components of Mindreading
The term mindreading has proven a useful banner to rally research efforts. But more work needs to be done to identify the basis functions underlying mindreading. In the language of the theoretician’s quadrant, to move forward, we must engage in a Q3 exercise.
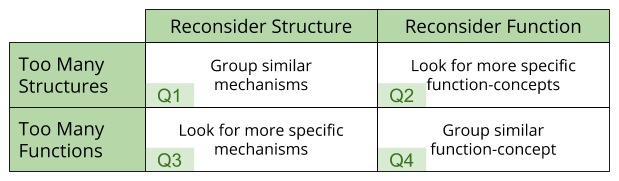
We have begun to sketch an outline of these basis functions during previous discussions of social phenomena.
- Social Behavior Network is an ancient faculty underlying sexual behavior, aggression, and parental care.
- Social Engagement System is an expression of polyvagal theory that explains how our facial musculature was recruited for social signaling.
- Language vs Communication explains how language is overlaid on top of the non-linguistic social engagement system.
- Argumentative Theory of Reasoning conceptualizes language as a more efficient vehicle for persuasion/evaluation and justification/evaluation.
- Gullible by Default explores the evaluation of arguments, showing how the ability to reject claims is a late addition to our cognitive toolkit.
- Counterfactual Simulation: Similar Until Proven Different explores the counterfactual mechanics of pretense, and how mindreading heavily relies on simulation.
- Agent Detection describes how agent detection relies on several more concrete faculties, such as face detection, movement detectors, and agent files.
- Intro to Goal Mindreading describes how equifinality might inform our ability to detect goals.
- Granite In Every Soul explores how we might employ agent files to conduct impression management.
- The Evolution of Faith discusses our species’ willingness to privilege social learning over individual learning.
Other mindreading-impacting phenomena we have not yet discussed include:
- Agency Detection. When the natural world violates our expectations (a leaf moves against gravity), often these events are caused by an (presently unseen) agent. Mismatches between agent and agency detection are thought to generate the intuitions that underlie our species’ folk animism.
- Emotion Contagion.
- Friendship behaviors.
- Shared Attention mechanisms. Before we can reason about the beliefs of another agent, we must learn to
- Cultural Psychological mechanisms. We have yet to discuss prestige biases, and our compulsive need to share information.
- Six Pillars of Selfhood. Kahneman distinguishes the Remembering vs Experiencing self. There’s
The following graphic attempts to bring together these subcomponents into a 10,000 foot view of the system. For more on this train of thought, I recommend Schaafsma et al (2015).
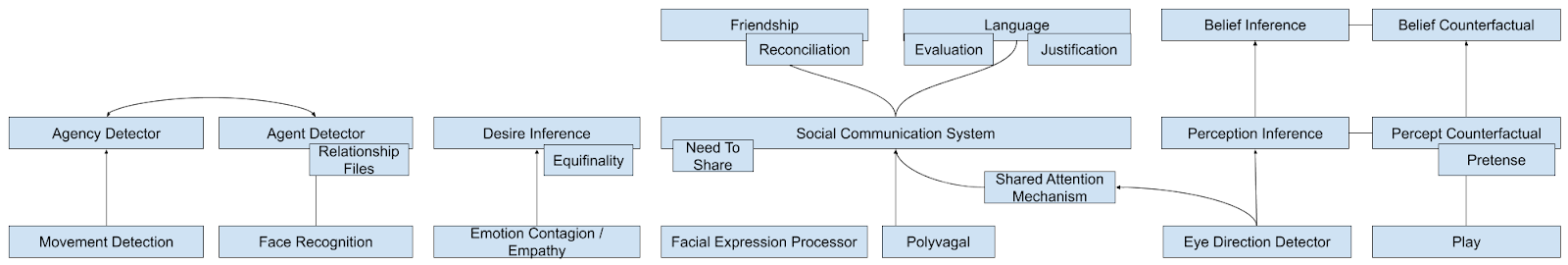
Clearly, representation mindreading involves more than a single faculty, but rather deploys a broad coalition of social faculties. It is likely that distinct “mindreading tasks” employed in experiments typically recruit coalitions with subtly different profiles.
Relationship to ICNs
The cognitive neuroscience community has converged on a set of neural mechanisms underlying goal and representation mindreading. These five regions are:
- Medial Prefrontal Cortex (MPFC)
- Posterior Cingulate Cortex (PCC)
- Temporo-Parietal Junction (TPJ)
- Superior Temporal Sulcus (STS).
- Temporal Pole (TP).
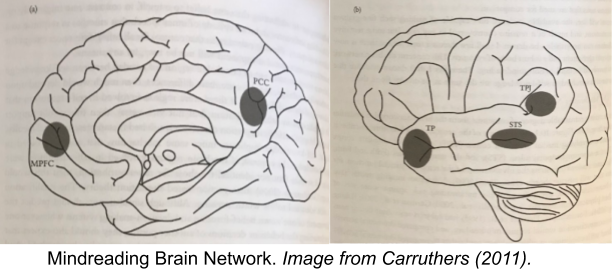
We have begun localizing specific functions to these five regions-of-interest. The TPJ seems to be the key site for representation mindreading, whereas goal mindreading is produced by the other sites; with the temporal pole appearing to underlie desire attribution specifically.
Scientific consensus is hard to achieve without a deluge of data; this network is here to stay. But there are two reasons to hesitate before drawing further conclusions. First, “mindreading” is probably not a natural kind; neural mechanisms probably map to more granular functions that join together to produce both macrosystems.
Second, these five regions must be structurally understood in terms of intrinsic connectivity networks (ICNs), and this work has not yet been undertaken. In my writeup of ICNs, we described evidence for five “processing networks”:
- Default mode network (and its three subcomponents)
- Salience Network and the closely related Ventral Attention Network (VAN)
- Dorsal Attention Network (DAN)
- Fronto-Parietal Control Network (FPCN) implicated in volitional control and willpower
- Cingulo-Opercular Control Network (COCN), implicated in working memory rehearsal and fluid intelligence.
The five regions of interest above are a subset of what social cognition theorists describe as the sociality network. In turn, the sociality network seems to comprise a subset of the default mode network. An increasing number of theorists are gesturing towards three subnetworks within the DM network, with mindreading modules mostly but not entirely residing within one of those subnetworks. Further, we have evidence that the default mode network is the basis of interoception and allostasis (that is, the brain’s unconscious representation of the body aka the hot loop).
These hints are suggestive. But precious little of our knowledge is detailed enough to be formalized and modeled. Someday I will be able to say more about the relationship between sociality, mindreading, interoception, and the default mode network. But that is not yet possible in 2020… at least, as far as I know.
Until next time.
References
- Baren-Cohen et al (1985) Does the autistic child have a “theory of mind”?
- Call & Tomasello (2008). Does the chimpanzee have a theory of mind? 30 years later
- Tomasello (2014). A Natural History of Human Thinking
- Gergely et al (1994). Taking the intentional stance at 12 months of age
- Heider & Simmel (1944) An experimental study of apparent behavior
- Schaafsma et al (2015). Deconstructing and reconstructing theory of mind
