
Part Of: Anthropogeny sequence
Followup To: An Introduction to Domestication
Content Summary: 2000 words, 20 min read
Two Forms of Aggression
Aggression is not a natural kind. Rather, as described in e.g., Siegel & Victoroff (2009), there are two kinds of aggression.
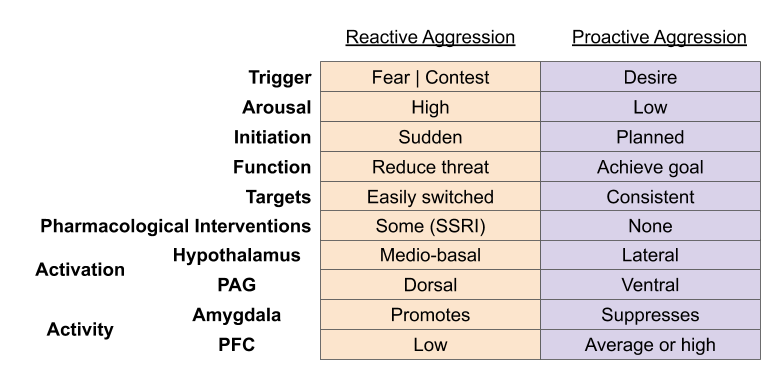
- Reactive aggression is based on the RAGE subsystem. It is the biological basis of resource competition.
- Proactive aggression is based on the SEEKING subsystem. It is the biological basis of predation, and sexual selection-driven infanticide.
These two systems have different behavioral signatures. Reactive aggression is associated with high arousal, sudden initiation, and functions to remove a threatening stimulus. As observers of a bar fight can tell you, you don’t want to get close to an enraged person at the wrong time – the aggressive behavior can easily switch its target. In contrast, proactive aggression is associated with low arousal, planned initiation, and functions to achieve some sort of goal.
These systems also feature different physiological signatures. Reactive aggression is caused by activation of the mediobasal hypothalamic nuclei, and dorsal nuclei of the periaqueductal gray (PAG). Amygdala activity promotes these behaviors, and are accompanied by low levels of prefrontal control. In contrast, proactive aggression is caused by activation of the lateral hypothalamic nucleus, and ventral regions of the PAG; amygdala activity suppresses its expression, and it is accompanied by significant cortical activity.
Of course, these two systems can interact.
- When a beta chimpanzee challenges an alpha, he may convert predatory aggression (plotting a coup) to an escalating sequence of reactive violence.
- When a human being suffers intense personal injury and is unable to immediately retaliate, he may convert that reactive rage into the more proactive and delayed phenomena known as vengeance.
The distinction also prominently appears in human legal codes: we tend to punish proactive aggression (premeditated murder) more virulent than reactive aggressive (bar fight).
Anyone looking at homicide data will tell you that being male, and being young, render a person much more likely to kill. Violence-generating mechanisms differ by sex, because each sex is subject to diverging selective pressures.
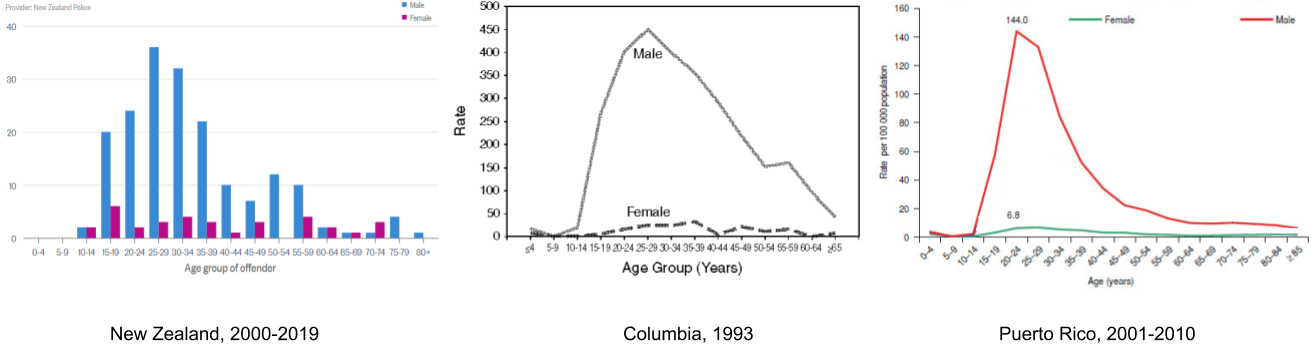
Of course, homicide can be produced by two kinds of aggression. It would be more useful to policy makers to analyze rates of reactive versus proactive aggression separately. Given its more cognitive basis, I suspect proactive violence is more amenable to cultural interventions; whereas reactive violence might be best treated with therapy and pharmaceuticals to strengthen one’s self-control.
And indeed, just these kinds of considerations are now being employed by social scientists seeking to better understand and mitigate phenomena such as domestic violence, and delinquency in children.

From a historical perspective, our species spent most of its history as foragers (i.e.,, hunter-gatherers), with statecraft a consequence of the agricultural revolution. There is a keen interest in understanding the natural tendency of forager populations, since these are more representative of the “original social contract”. The Rousseau paradigm sees foraging humans as a naturally benign and unaggressive species. This position considers violence to be promoted by the state. The Hobbes paradigm rejects the idea of the noble savage and holds violence in the evolutionary path. In this view, the state is an instrument to restrain violence.
The Evolution of War
Comparative biology data can resolve the Rousseau-Hobbes debate.
First, consider how chimpanzees use gangs of allied individuals to achieve political ends through aggressive means. These coalitions are very rare in the animal kingdom. They are only known to occur among social carnivores and primates. These acts of coalition-based aggression are proactive in nature.
Second, it is important to understand how chimpanzees express xenophobia. Chimp troops don’t wander haphazardly; they instead inhabit clearly demarcated territories. The troops of neighboring communities are treated with hostility, so much so that up to 75% of the time is spent in the central 35% of the range. Another expression of chimp territoriality is border patrols, conducted by groups of male chimps moving stealthily to enforce their territory’s boundaries.
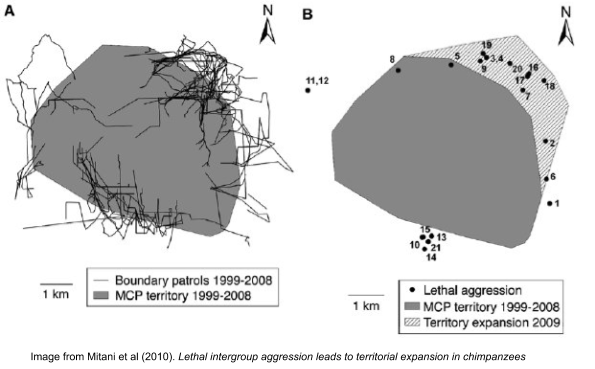
Third, these factors coalesce in the phenomenon of chimpanzee commando raids, with large groups of males penetrating deep into enemy territory, stalking and killing members of competing troops. Why small-scale raids instead of large-scale brawls? Well, warfare is only adaptive when the potential benefits outweigh the risk of personal injury. Thus, these raids are governed by the logic of a local imbalance of power. Raids preferentially occur when the attacking party has gathered significantly more fighting power than the defender (Wrangham 1999).
Killing doesn’t directly increase one’s biological fitness. Why then has such behavior been selected? Because successful raids promote the possibility of territorial expansion (Mitani et al 2010), plausibly by weakening the other groups’ overall fighting power. In turn, territory size directly correlates with resource and mate availability.
Here is Wrangham (1999) explaining parallels with human warfare:
It is clear that intergroup aggression has occurred among many, possibly all, hunter-gatherer populations and follows a rather uniform pattern. From the most northern to the most southern latitudes, the most common pattern of intergroup aggression was for a party of men from one group to launch a surprise attack in circumstances in which the attackers were unlikely to be harmed. Attacks were sometimes unsuccessful but were, at other times, responsible for the deaths of one or many victims. Women and girls were sometimes captured.
Chimpanzees and hunter gatherers, we conclude, share a tendency to respond aggressively in encounters with members of other social groups; to avoid intensely aggressive confrontations in battle line (typically, by retreating); and to seek, or take advantage of, opportunities to use imbalances of power for males to kill members of neighboring groups.
Indeed, even the rate at which foraging humans and chimpanzees engage in between-group violence is quite similar:
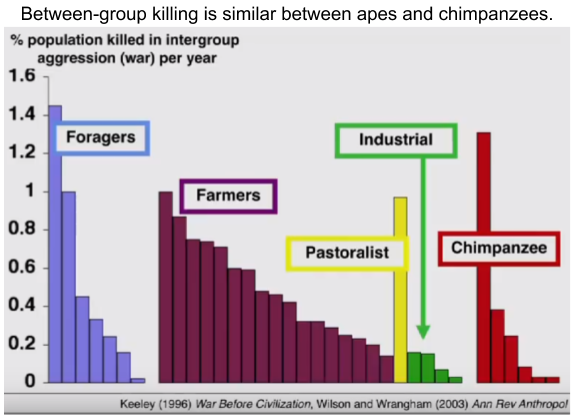
These data suggest a common mechanism. It is not that humans evolved a unique thirst for warfare. Rather, this instinct long predates our species.
The Domestication of Bonobos
It is hard to imagine species with more dramatically different social lives than bonobos and chimps. They are renowned for their ultra-sexualtity: sexual acts are used in lieu of grooming, as the primary vehicle to strengthen relationships. They also exhibit startlingly low rates of violence:
- Killing of any kind (including coalition-based acts of violence) is literally unheard of.
- Rape and infanticide have also never occurred.
- Commando raids do not occur; bonobos do not even express hostility to neighboring troop “outgroups”.
The bonobos and chimp lineages diverged very recently (less than 1 mya); yet they lead entirely different social lives. How is this possible?
A clue comes from observations of unusually strong female coalitions in bonobos. Every time a male tries to coerce a female for food or sex, that female’s coalition vigorously rebuff the coalition. These female coalitions in effect give non-aggressive males an advantage. Over the generations, this selective pressure will yield decreasing levels of (proactive) aggression in the bonobo species.
As we learned in An Introduction to Domestication: when aggression is downregulated in a species, a whole complex of unintended byproducts occur. And we see precisely this domestication syndrome in bonobos. Bonobos have smaller crania, reduced pigmentation, increased sexual behaviors, and a general uptick in childlike mannerisms. Bonobos domesticated themselves! Here is the model from Hare et al (2012):
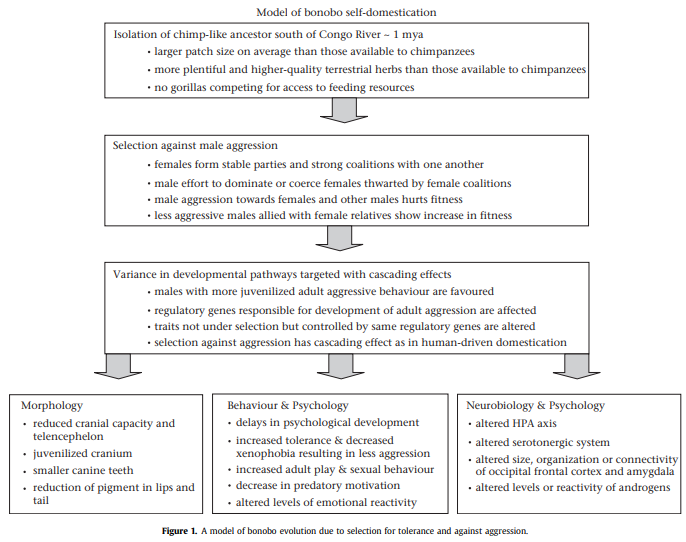
The Puzzle of Humanity
Chimpanzees and humans have comparably high rates of proactive (predatory) violence, and this proclivity underlies a shared love of warfare. In contrast, bonobos exhibit near-zero rates of proactive aggression.
Let’s turn our attention back to reactive violence. Bonobos exhibit moderate forms of reactive aggression; primarily expressed by female coalitions to curtail male domination behaviors. In contrast, chimps are notoriously short-tempered; reacting violently to even trivial “provocations”. How do rates of human reactive aggression compare in practice?
Even in comparatively violent forager groups, the difference is remarkably large. Humans experience reactive violence at rates two orders of magnitude less than our chimpanzee cousins.
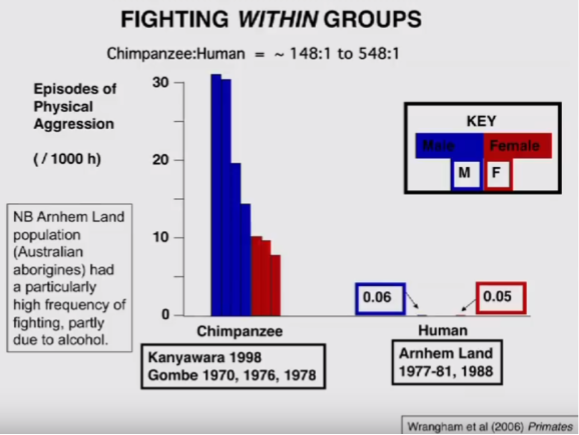
With these data, the following picture has emerged:
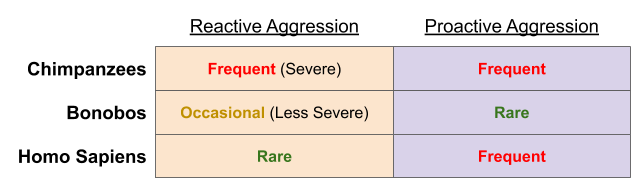
Let’s assume chimpanzee aggression behaviors are representative of the LCA. Bonobos became docile by a process of self-domestication. Why are humans less reactively violent? Did we self-domesticate too?
Another Case of Self-Domestication
The surprising answer is yes. A host of anatomical changes in H. Sapiens around 300 ka all support the self-domestication hypothesis (Leach 2003, Cieri et al 2014).
One symptom of domestication is paedomorphism: childlike features that extend into adulthood. Our adult cranium (especially the smooth, round skull) resembles the skull of chimpanzee children (in contrast with a chimpanzee adult’s prognathic face):

In domestication (among others), we see a reduction in face size, and a feminization of the skull:
These changes look a lot like the change between our mid-Pleistocene ancestor and anatomically modern H. Sapiens:
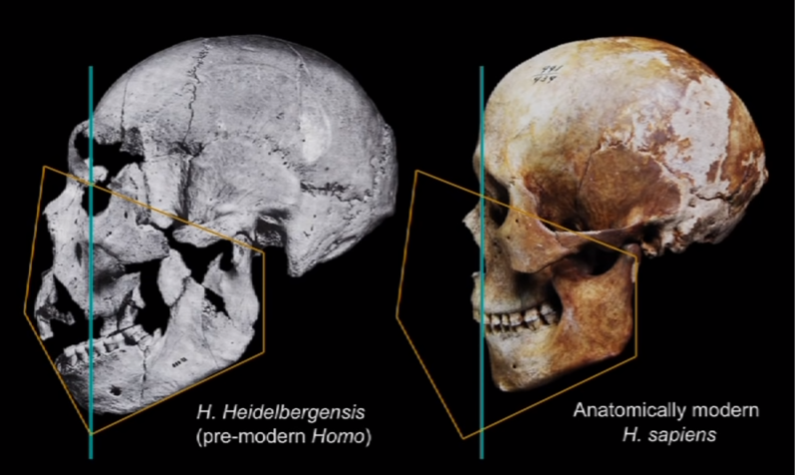
Other “domestication signatures” in modern humans include:
- Brain volume reduction (in last 30,000 years)
- Smaller teeth, small face-body ratio
- Reduced sexual dimorphism (differences between male vs female skeletons)
- More childlike features in adults (longer juvenile period, extended learning, adult play)
- Increased fertility rate (incl. hidden estrus)
- Increased rates of lifelong homosexuality
This anatomical evidence of self-domestication nicely explains with our species’ unique relationship with violence.
Significance of Self-Domestication
Consider again that the domestication syndrome appears between 400 and 100 kya. The Heidelbergs were more violent than Sapiens.
Most primates don’t get enraged by acts of violence that don’t involve them personally. But humans do experience moral outrage at such acts, to the point of being willing to engage in so-called altruistic punishment (risking personal injury to punish a third-party offense).
Moral instincts are one of the couple dozen traits that are uniquely human. Evolutionary anthropology must explain when and why these uniquely human faculties were forged. Being willing to punish acts of reactive violence surely played a role in the self-domestication process. We can safely conclude that morality as a cognitive adaptation evolved late. Heidelbergs were amoral; Sapiens were increasingly subject to the moral sentiments.

I’ll speak towards why morality evolved another time. For now, let’s turn our attention from the causes, to the effects of self-domestication. For it turns out that these data give us unique insights into what made our species ecologically dominant. Heidelbergs did not conquer the globe – Sapiens did. But how could a reduction in intra-group violence create the necessary conditions for our species’ success?
Our species was not successful because of its pacifism. Rather, the cultural intelligence hypothesis holds that our species’ unique gifts for coordinating with others to transmit cultural knowledge created the conditions for cultural ratcheting. Rather than inheriting only our genetic legacy, we also inherit cultural knowledge which (together with our innate endowments) give us increasing powers to control our environment.
Importantly, the ability for our cultural knowledge (or “super mind”) to accumulate information is not guaranteed. If a particular community of humans is too few in number, or too antagonistic towards one another, its net cultural know-how will not grow across the generations.
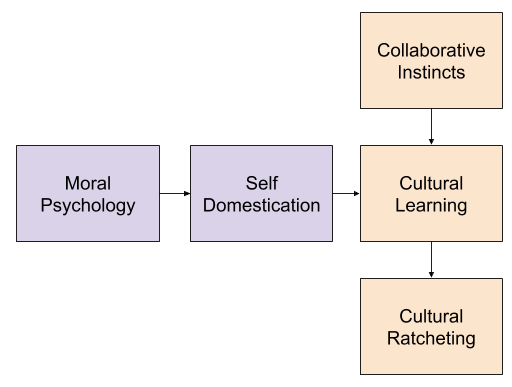
In this model, our cultural instincts evolved earlier in our lineage. But the advent of morality, and its concomitant reduction in reactive violence, was the event that unleashed the astonishing generative potential of human culture.
Until next time.
Inspiring Materials
Some of these views are articulated in more detail in Wrangham (2019a). For video lecture on this topic, please see:
Works Cited
- Cieri et al (2014). Craniofacial Feminization, Social Tolerance, and the Origins of Behavioral Modernity
- Hare et al (2012). The self-domestication hypothesis: evolution of bonobo psychology is due to selection against aggression
- Leach (2003). Human Domestication Reconsidered.
- Marean (2015). An Evolutionary Anthropological Perspective on Modern Human Origins
- Mitani et al (2010). Lethal intergroup aggression leads to territorial expansion in chimpanzees
- Siegel & Victoroff (2009). Understanding human aggression: New insights from neuroscience
- Wrangham (1999). Evolution of Coalitionary Killing
- Wrangham (2003). Intergroup Relations in Chimpanzees
- Wrangham (2018). Two types of aggression in human evolution
- Wrangham (2019a). The Goodness Paradox: The Strange Relationship Between Virtue and Violence
- Wrangham (2019b). Hypotheses for the Evolution of Reduced Reactive Aggression in the Context of Human Self-Domestication
