
Part Of: Biology sequence
Followup To: The Evolution of Senescence
Content Summary: 2400 words, 12 min read
The Fast-Slow Continuum
Across the life course, an organism must choose between somatic investment (growth, learning, maintenance) vs reproductive effort (parenting, alloparenting, mating). But there are also trade-offs within each of these investment categories. For example, within reproductive effort, should I emphasize mating or parenting?
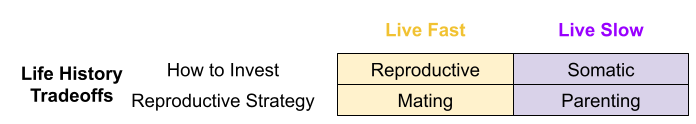
We can reformulate these tradeoffs as current vs future reproduction, and offspring quantity vs quality. Due to sexual selection asymmetries, first trade-off tends to be more salient to females, and the second is more salient to males.
The effect of these tradeoffs can be detected in various life history (LH) traits:
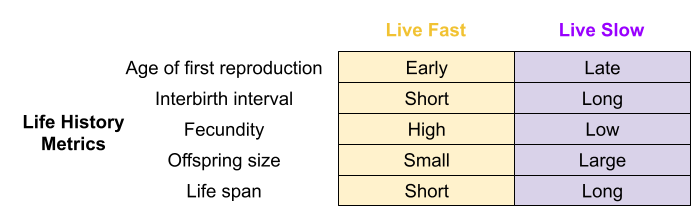
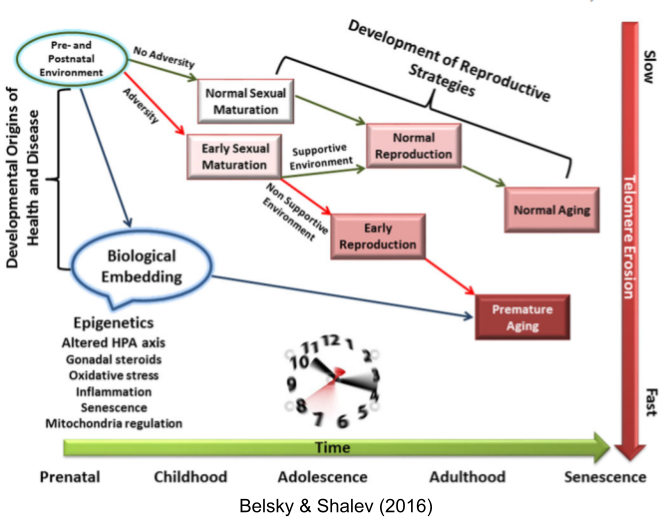
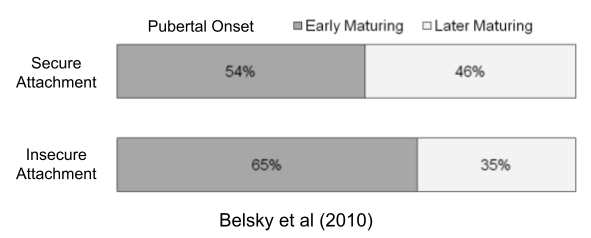
The disposable soma theory posits a causal chain: higher environmental mortality → prioritized reproductive effort → increased senescence. But faster aging also correlates with early puberty (Belsky & Shalev 2016). Perhaps both LH traits respond to the same life-history tradeoff?
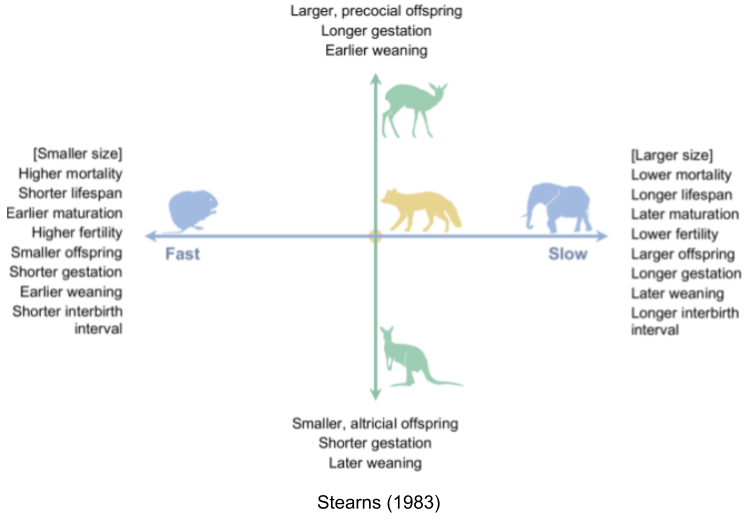
If you compare all five life history traits from species-typical traits across all species, they all run together (Jeschke et al 2008).
- Fast species show early reproductive maturity and short lives. Semelparous species like salmon undergo “programmed death” immediately after reproduction.
- Slow species live longer, and take more time to reach reproductive maturity. For example, iteroparous species like blue whales have extremely slow lifespans.

This is the fast-slow continuum. This is supported by comparative analyses in mammals (Bielby et al. 2007), birds (Saether 1988), reptiles (Bauwens & Diaz-Uriarte 1997), fish (Winemiller & Rose 1992), insects (Johansson 2000) and plants (Salguero-Gómez et al. 2016).
In fact, two components stand out in dimensionality reduction analyses. A fast-slow axis that accounts for 70-80% of the variance in the traits, and a precocial-altricial axis that explains 10-15% of the variance. While smaller species tend to be fast and vice versa, controlling for body size does not make the continuum disappear; rather, the respective variances change to 30-50% and 20-30% (Del Guidice 2020).
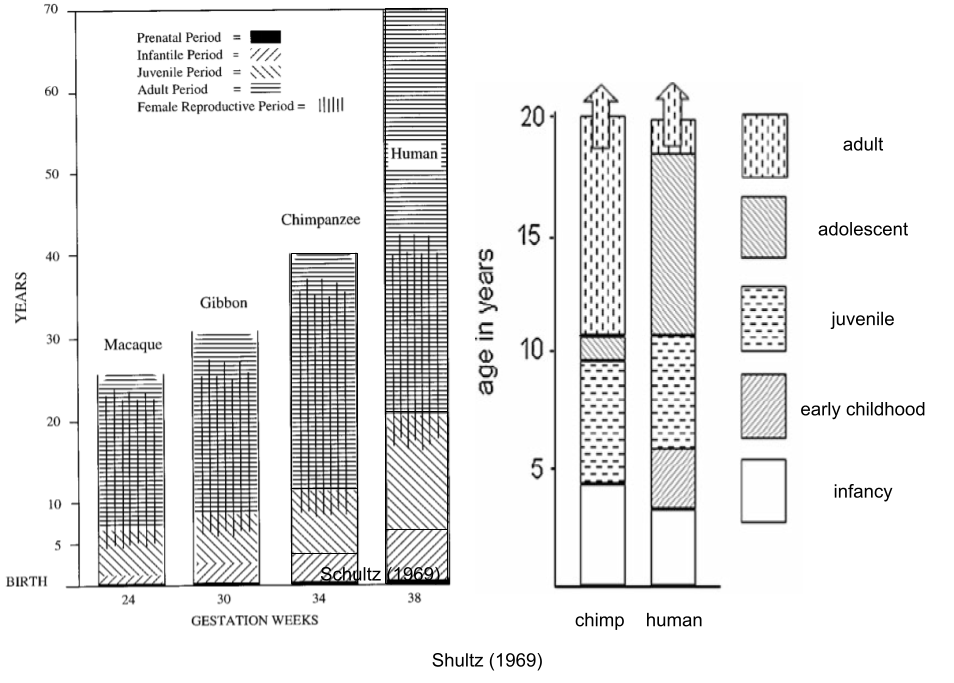
As an aside, our species presents a kind of exception to the fast-slow continuum! Human longevity is more “slow” than chimpanzees. But we also have a shorter interbirth interval, a “fast” characteristic. This energetic paradox is explained by a lifting of energetic constraints (somatic vs reproductive effort – why not both?). It was funded by an increase in human basal metabolic rate (Pontzer et al 2016); this expanded energy budget funded both increased reproductive output (some ~100 kcal per day) and somatic costs (larger brains, longer lifespans, etc).
Explaining The Continuum
The fast-slow continuum is a reliable empirical pattern. Why does it exist? What causes these interspecies differences in the pace of life (POL)?
In the 1970s, three US ecologists proposed the r/K theory of life history, which strove to explain it by appealing to population density and carrying capacity of the environment. But by the 1980s it was clear that carrying capacity could not explain the continuum.
Three theoretical approaches stand out:
- Modern density dependent models (e.g., Engen & Sæther 2016) do away with carrying capacity, but extend classical r/K density concepts.
- Allometric models (e.g., the metabolic theory of ecology in Brown et al 2004) can explain body size covarying with life history; but struggle to predict the continuum’s persistence after body size is partialed out.
- Other models invoke extrinsic mortality (risk beyond an individual’s control). Extrinsic mortality affects LH traits only through modifying the intensity of competition: higher extrinsic mortality reduces the intensity of competition and vice versa (Andre & Rousset 2020).
But I haven’t yet encountered a “theory of everything” that integrates across these perspectives. Data in quantitative ecology tends to be rather sparse, making generalizations difficult.
Personality & Plasticity
Let’s talk about animal personality. Numerous studies have found that some individuals are consistently bolder (Wilson et al 1994) or more aggressive (Johnson & Sih 2007) than others across multiple situations, and indeed that boldness and aggressiveness are often positively correlated (Bell & Sih 2007). Such behavioral syndromes have been seen in a wide range of taxa (Gosling 2001).
Why should animals have a personality as opposed to being completely flexible? Why are human personalities so pervasive that when someone is a behavioral chameleon, we often view them as psychopathic?
Behavior could be, in principle, completely flexible (i.e, an animal could be highly aggressive one moment, and then cautious shortly thereafter). But if optimal behavior is connected to a slow-changing state variable, then adaptive behavior should also be consistent over long periods. A few examples from Sih (2011):
Personality is thought to emerge from feedback loops between state and behavior (Sih et al 2015). Negative feedback tends to remove individual differences; positive feedback tends to accentuate them. A few more examples:
Individual differences are often grounded in genetics. For example, twin studies models suggest that 50-80% of the variance from pubertal onset is genetic (Rowe 2002). But the above data suggests a role for environmental influence. We must therefore model organism plasticity, or conditional adaptations in response to environmental conditions.
The Pace of Life Syndrome
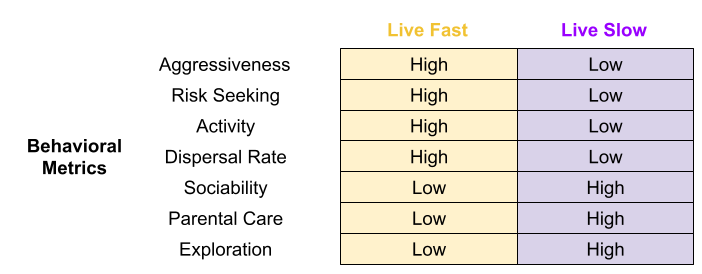
Behavioral syndromes within species bear a striking resemblance to interspecific fast-slow differences. Links have been found:
- … between aggressive, risky behaviors and high reproductive success but also to lower survival (Smith & Blumstein 2008). Found in bighorn sheep (Reale et al 2009) and red squirrels (Boon et al 2008).
- … between aggressive individuals and dispersal (Dingemanse et al. 2003).
- … between aggressive males and less parental care (Duckworth & Badyaev 2007).
- … between sociability and reduced propensity to disperse (Cote et al 2010).
Perhaps the conditions that generate interspecific POLS also generate interindividual POLS. This working assumption is known as the ecological gambit. The gambit becomes riskier if causal factors are known to operate at one level of analysis, but not another (Pollet et al., 2014). But the assumption is not without support. Some density dependent models that explain species POLS concurrently explain individual differences (Wright et al 2019). Finally, the Andre & Rousset (2020) model shows that extrinsic mortality influences individual, not just species, differences in life histories.
We can visualize the relationship between species and individual-level pace of life with the following cartoon. The biggest dark-blue oval represents the entire fast-slow continuum, the smaller blue ovals each represent a species, and the yellow ovals represent communities or individuals within a given species.
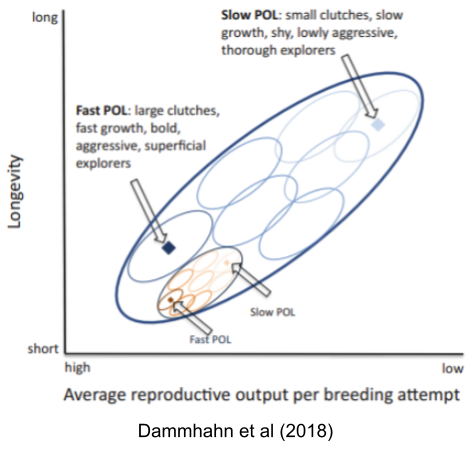
Behavior co-evolves with anatomy. If an individual’s body decays more quickly (preferentially investing resources into growth and reproductive effort), we should expect that individual to adopt different behaviors. Fast bodies should exhibit behaviors such as aggressiveness (especially in the context of intrasexual contest competition), and risk seeking. In fact, many behavioral metrics arguably conform to the fast-slow continuum.
Most sections in this article are fairly uncontroversial. However, the topics of this section are more contentious. Not everyone agrees that POLS can be applied to individuals and behaviors; some prefer to keep the analysis to average traits across species.
Evaluating the Syndrome
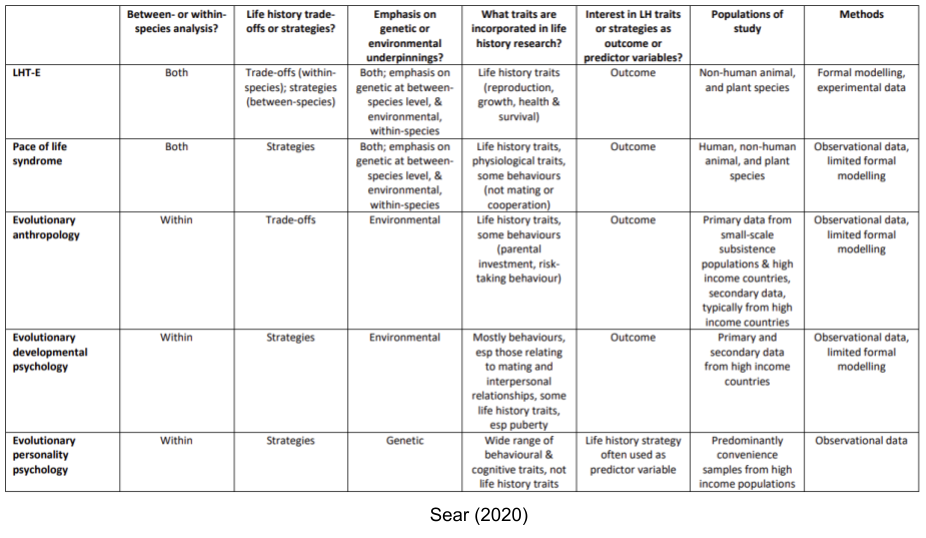
Life history theory (LHT) can be usefully understood as two sub-disciplines, whose literatures operate largely independently of one another (Nettle and Frankenhuis 2019)
- LHT-E, or life history theory as practiced in ecology and evolutionary biology.
- LHT-P, or life history theory as practiced in evolutionary psychology.
LHT-E tends to focus on the POLS traits across species. Within LHT-P, there are four separate movements, with decreasing levels of contact with evolutionary biology: pace of life syndrome research, evolutionary anthropology, evolutionary developmental psychology, and evolutionary personality psychology.
One of the strongest criticisms of LHT-P is its reliance on verbal models in lieu of formal models. As argued by Stearns & Rodrigues (2020), our capacity to develop complex verbal arguments is limited (the chamber of consciousness is small!). Our intuition often misleads, and the sensitivity of predictions to their underlying assumptions is not easy to see within informal arguments.
For example, Darwin’s several attempts to explain verbally the evolution of 50:50 sex ratios were unsuccessful: “I now see that the whole problem is so intricate that it is safer to leave its solution for the future” (Darwin 1874, pp. 259, 260). This problem was later solved in a few lines of algebra (Edwards 1998, 2000).
Another important criticism is a reliance on questionable assumptions of “extrinsic mortality”, which too often conflates random vs condition-dependent forms of environmental mortality.
The fast-slow continuum seems to be settled science. But animal personality research in general, and POLS in particular, is on empirically shaky ground (Mathot and Frankenhuis 2018). Yet I suspect more conservative examples of LHT-P are worth attending to.
But the construct seems to explain a great deal of physiology. Chronic stress may not be a disease; it may canalize the fast phenotype.
Chronic Stress vs. Pubertal Onset
The development of individual organisms is subject to ecological factors that drive interspecific differences. Human females, for example, can experience rapid (k1) or slow (k4) somatic growth. Each growth curve corresponds to a different optimal age of reproductive maturity; the solid line denotes this reaction norm.
Compared to the 19th century, women have become younger and taller at age of maturity (Worthman 1999). This secular trend has been most intense within groups of low socioeconomic status (SES) (e.g., Abioye-Kuteyi et al 1997). This class effect only pertains to countries where low SES groups do not suffer from systematic malnutrition and disease (it is absent in e.g., Denmark Helm & Lidegaard 1989). Interestingly, the link between nutrition and earlier puberty may be partially mediated by fiber content in the diet (Koo et al 2002). Other physical stressors can delay female puberty. While some exercise promotes androgens (Elias 1981), professional athletes often experience delayed puberty (e.g., Bale et al 1996).
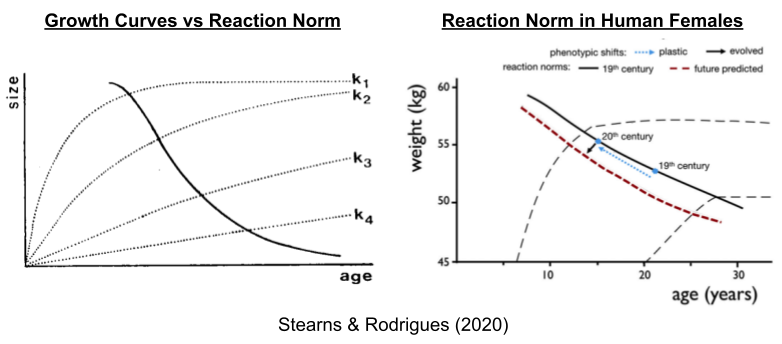
Taken together, the shift up the reaction norm occurred with the Industrial Revolution and its concomitant energy surplus. Higher activity in the HPG axis is associated with earlier puberty; which may explain why levels of testosterone are also higher in developed countries (Henrich 2020 pp 551).
But some stressors can have the opposite effects. Survivors of the Great Sichuan earthquake were twice as likely to experience early puberty than controls (Lian et al 2018; see also Pesonen et al 2008). And more generally, a substantial literature indicates that early exposures to childhood adversities (e.g., socioeconomic adversity, childhood attachment, heightened parent–child conflict, father absence) tend to predict earlier pubertal development in females (Ellis 2004).
How to reconcile these findings? It seems like pubertal timing is contingent firstly on health and nutrition (see especially Kyweluk et al. 2018) and secondly, when these are adequate, on socioemotional conditions (Coall & Chisholm 2003).
Biological Substrate
Recall the big life history tradeoffs faced by organisms:
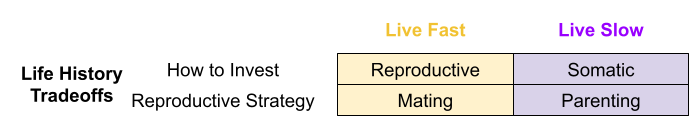
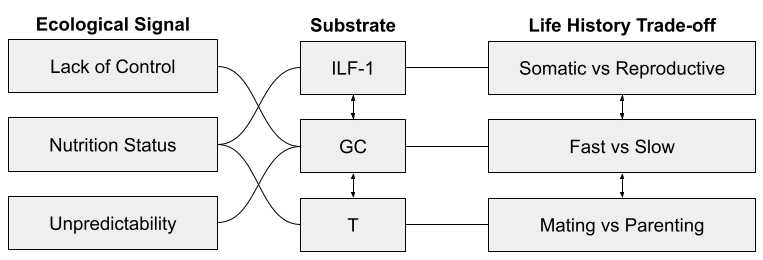
Three endocrine systems are involved in these dimensions:
- I’ve previously noted how insulin-like growth-factor 1 (IGF-1) seems involved in the tradeoff between somatic and reproductive effort. It is especially responsive to nutritional status.
- I have elsewhere noted how glucocorticoids (GC), and the stress response system (SRS) more generally, seem to mediate fast and slow phenotypes. The SRS is especially responsive to unpredictability and lack of control.
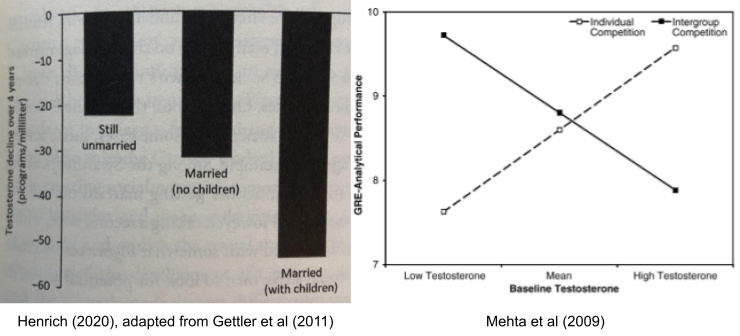
- Finally, testosterone (T) plays a role in mating vs parenting decisions, at least in males. Testosterone drops substantially after marriage, and again on the birth of a child. It also appears to promote intragroup competition, consistent with sexual selection theory:
It may be worth exploring interactions between these different endocrine systems as they unfold across development.
Until next time.
References
- Abioye-Kuteyi et al (1997). The influence of socioeconomic and nutritional status on menarche in Nigerian school girls
- Andre & Rousset (2020). Does extrinsic mortality accelerate the pace of life? A barebones approach
- Baams et al (2015). Transitions in body and behavior: a meta-analytic study on the relationship between pubertal development and adolescent sexual behavior
- Bale et al (1996). Gymnasts, distance runners, anorexics body composition and menstrual status.
- Belsky et al (2010). Infant attachment security and the timing of puberty: testing an evolutionary hypothesis
- Belsky & Shalev (2016). Contextual adversity, telomere erosion, pubertal development, and health: two models of accelerated aging, or one?
- Bergmuller et al (2010). Animal personality due to social niche specialization
- Boon et al (2008). Personality, habitat use, and their consequences for survival in North American red squirrels (Tamiasciurus hudsonicus).
- Brown et al (2004). Toward a metabolic theory of ecology
- Clark (1994). Antipredator behavior and the asset protection principle.
- Cote et al (2010). Social personalities influence natal dispersal in a lizard.
- Dall et al (2012). An evolutionary ecology of individual differences.
- Dammhahn et al (2018)
- Del Guidice (2020). Rethinking the Fast-Slow Continuum of Individual Differences
- Dingemanse et al (2003) Natal dispersal and personalities in great tits (Parus major).
- Duckworth & Badyaev (2007). Coupling of dispersal and aggression facilitates the rapid range expansion of a passerine bird.
- Elias (1981). Serum cortisol, testosterone, and testosterone-binding globulin responses to competitive fighting in human males
- Ellis (2004). Timing of pubertal maturation in girls: an integrated life history approach
- Engen & Sæther (2016). Optimal age of maturity in fluctuating environments under r- and K-selection
- Gettler et al (2011). Cortisol and Testosterone in Filipino Young Adult Men: Evidence for Co-regulation of Both Hormones by Fatherhood and Relationship Status
- Helm & Lidegaard (1989). The relationship between menarche and sexual, contraceptive, and reproductive life events
- Henrich (2020). The WEIRDest people in the world: how the West became psychologically peculiar and particularly prosperous
- Jeschke et al (2008) r-Strategist/K-strategists.
- Koo et al (2007). A cohort study of dietary fibre intake and menarche
- Kyweluk et al (2018). Menarcheal timing is accelerated by favorable nutrition but unrelated to developmental cues of mortality or familial instability in Cebu, Philippines.
- Lian et al (2020). The impact of the Wenchuan earthquake on early puberty: a natural experiment
- Mathot and Frankenhuis (2018). Models of pace-of-life syndromes (POLS): a systematic review
- Mehta et al (2009). When are low testosterone levels advantageous? The moderating role of individual versus intergroup competition
- Nettle & Frankenhuis (2020). Life-history theory in psychology and evolutionary biology: one research programme or two?
- Pollet et al (2014). What can cross-cultural correlations teach us about human nature?
- Pontzer et al (2016). Metabolic acceleration and the evolution of human brain size and life history
- Pesonen et al (2008). Reproductive traits following a parent-child separation trauma during childhood: a natural experiment during World War II.
- Reale et al (2009). Male personality, life-history strategies and reproductive success in a promiscuous mammal
- Reale et al (2010). Personality and the emergence of the pace-of-life syndrome concept at the population level
- Sear (2020). Do human ‘life history strategies’ exist?
- Stearns & Rodrigues (2020).
- Shultz (1969). The life of primates
- Smith & Blumstein (2008). Fitness consequences of personality: a metanalysis.
- Stearns (1983). The influence of size and phylogeny on patterns of covariation among life-history traits in the mammals.
- Worthman (1999). Evolutionary perspectives on the onset of puberty.
- Wright et al (2019). Life-history evolution under fluctuating density-dependent selection and the adaptive alignment of pace-of-life syndromes
