
Part Of: Anthropogeny sequence
Followup To: Intro to Multilevel Societies
Content Summary: 2400 words, 12 min read
Introduction
In today’s post, I will argue that humans have a multi-level society (MLS) social organization, and also evolved as cooperative breeders (CB). These attributes together explain a great deal of human uniqueness. They are an explanatory bridge over the yawning chasm, from the other great apes to our species
MLS+CB animals are very rare in mammals, but common in birds. I will also argue our ancestors took this avian-like pathway with the advent of early menopause.
Multilevel Societies in Humans
Social organization in animals can be categorized into four distinct types.
- Solitary Living: Every individual has its own territorial stake on foraging real estate, and guards access. Solitary females will reproduce with solitary males at the boundary of its territory, but raise the child alone. The pair-bond is exclusively expressed in the relationship between mother and her children.
- Monogamous Pair-Living.Female and male animals will co-reside in the same territory. The pair-bond is extended to support not just parenthood, but also a romantic commitment between the reproducing parents.
- One-Male Units (OMU). Adult males engage in contest competition for women which ultimately take the form of one-male multi-female harems. Sexual body dimorphism is accentuated due to the selective pressure of inter-male competition. Because of polygyny’s math problem, bachelor male primates often cause problems for the established “families”.
- Multi-male Multi-female (MM-MF) groups. Adult males don’t typically fight over access to women, sexual selection in these societies tends towards sperm competition instead. Female sexuality is promiscuous, which can be interpreted as a paternal uncertainty device.
To be extremely reductive, MM-MF groups possess groups, but lack families and love. Monogamous and OMU social structures have families and love, but lack groups.

Every animal species exhibits one of these social systems. Which one best characterizes Sapiens? Well, iIt is natural to identify as a family-living species. But pair-living and OMU animals don’t live in groups; our multifamily groups are anathema. What gives?
The discovery of multilevel societies (MLS) gives us the language to dramatically improve our understanding of human social organization. There are some species that exhibit a blend of both family- and group-living. From a phylogenetic perspective, MLS seem to emerge from two pathways:
- With the bonding pathway, pair bonding nucleation occurs within ancestral mm-mf groups.
- But with the aggregation pathway: autonomous OMUs increasingly overlap, and ultimately affiliate.

Enriched Male Coalitions in MLS
To quote from Chapais (2008)
It has been estimated that female chimpanzees copulate ~1500 times per conception. Consistent, long-term preferential relationships between males and females have not been observed. Paternal certainty is near zero, as is paternal investment. In such promiscuous systems, nearly all (95%) of siblings are half siblings.
Consider the perspective of a single male chimpanzee, Ego. Ego can identify who his mother and siblings are, by virtue of developmental familiarity. The reliable ability to recognize one’s siblings is the basis of the Westermarck effect.
But Ego faces two barriers to kin recognition. First, due to parental uncertainty in promiscuous MM-MF settings, a male chimpanzee cannot identify his father (nor, in consequence, any of his father’s relatives). He also cannot identify his own sons and daughters, nor the offspring of his brothers. Second, chimpanzee societies are organized by female dispersal, with females relocating to other troops when they reach reproductive age. Without this emigration, Ego could recognize his mothers relatives, and his sister’s children… but he will never meet them (male chimpanzees cannot visit neighboring communities, because they would be killed by resident males).
The theory of inclusive fitness predicts substantial cooperation between relatives. But kinship-based cooperation is viable only amongst kin you can recognize. So, male coalitions are very small in male philopatric societies.

But multilevel societies (MLS) reintroduce the pair bond, and paternal uncertainty is lessened. Ego is suddenly able to identify his father, and – by social inference – his father’s siblings. Ego is also able to recognize his brother’s offspring (but not his sister’s offspring, since she still disperses before reproducing).
In principle, Ego can also identify his son’s children, and his father’s parents — but in practice, chimpanzee life history is such that only three generations typically coexist (these possible recognitions are denoted in light green).

Kin recognition is expanded in MLS societies. The male cooperative networks have become much more powerful! In our example, rather than having one ally (his brother), Ego is able to forge alliances with five other males.
Male cooperative networks were further expanded by the ability to recognize affines (in anthropomorphic terms, “brother-in-law”), whose shared interest in their offspring provided another kinship basis for cooperation.
Reinterpreting Human Monogamy
Consider the following facts:
- Our closest ancestors are either MM-MF (chimpanzees) or OMUs (gorillas). None of them are monogamous pair-living.
- On average, male body size is about 15% larger than female body size. Sexual dimorphism of 15% is not what we would expect from a monogamous species.
- We know of zero examples of a monogamous species evolving an MLS organization. In contrast, OMUs and MM-MF species become MLS often.
- In monogamous primates, sexual coercion of females is rarely, if ever observed. Mate guarding and contest competition are also largely absent.
In fact, polygyny seems to be the cross-cultural human universal. Not monogamy.
For a given band of foragers, we can count how many men have one wife, and how many have more than one. Marlowe (2003) reports that polygamously-married men exist in some 90% of societies in the standard cross-cultural sample (SCCS). To be more specific, 60% of societies have about ~10% of men with more than one wife, and 30% of societies have even more sexual inequality than this.
Polygyny creates a “math problem”: it inevitably creates pools of unmarried, low-status males.
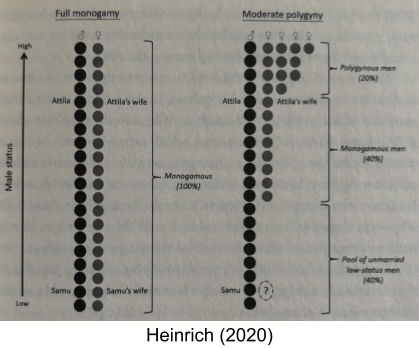
All polygynous societies have bachelor males (“incels”) who harass males in established harems. But in MLS societies, bachelor primates cooperate in all-male bands and try to wrest breeding opportunities from the “married” males (Qi et al 2017). This generates even more selective pressure from male cooperation in the breeding bands.
Henrich (2012) offers an explanation for the recent phenomenon of state-enforced monogamy.
Taking wives is always positively associated with status, wealth or nobility, even among highly egalitarian foraging societies. After the origins of agriculture, as human societies grew in size, complexity and inequality, levels of polygynous marriage intensified, reaching extremes in the earliest empires whose rulers assembled immense harems. Yet, monogamous marriage has spread across Europe, and more recently across the globe, even as absolute wealth differences have expanded. Here, we develop and explore the hypothesis that the norms and institutions that compose the modern package of monogamous marriage have been favored by cultural evolution because of their group-beneficial effects—promoting success in intergroup competition.
Intro to Cooperative Breeding
Eusocial insects, such as bees, are characterized by caste-based division of labor, cooperative raising of children (i.e., alloparenting), extreme reproductive skews (e.g., reproduction monopolized by the queen), and ultracooperative behaviors leading to colonies behaving as a cohesive superorganism.
E.O Wilson (1975) famously called humans eusocial apes, appealing to our own species’ ultracooperative behavior. Eusociality was once thought to exist only in insects, but striking evolutionary convergences have since convinced us of a eusociality continuum (Sherman et al 1995) between these insects and cooperatively breeding birds and mammals.
Most primates adopt a continuous care and contact (CCC) parenting model, with the infant maintaining an unceasing grasp on his mother’s fur until they reach maturity. But CCC is a last resort for primate mothers who lack safe alternatives. When the species-specific frequency of male infanticide is low (as it is for humans), maternal hyperpossessiveness is relaxed (Hrdy 2009).
In contrast with monkeys, mothering does not “come naturally” for great apes. Infant mortality for first time mothers is surprisingly high. Little surprise, then, that young female apes are drawn to practice mothering.
Compared to their non-cooperative breeding peers, species that do use alloparenting exhibit shorter interbirth intervals, which dramatically improves their ability to colonize new habitats. The family Callitrichidae, for example, are famous for breeding fast and for their rapid colonization of new habitats.
More interesting, species with alloparenting exhibit many more prosocial behaviors than those without it (Burkart & van Schaik 2010). These include enhanced mindreading abilities, shared intentionality, increased social tolerance, and spontaneous prosociality.
Why might explain the link between alloparenting and prosociality? Consider the plight of an infant raised in such an environment. Their survival is no longer the exclusive domain of their mother; but also depends on soliciting help from other alloparents – some of whom may not even be kin. Consequences for failure are large: unlike CCC, mothers in cooperative breeding societies exhibit high rates of child abandonment, when they assess insufficient resources are available.
For the infant, this sets off a kind of social Machiavellian environment. Infants must attend to the mental state of nearby conspecifics, and tune their solicitations accordingly. In order to “stay in touch without touch”, infants use vocalizations like crying and babbling. This may explain why shared intentionality develops as early as 9 months in human infants (Tomasello & Gonzalez-Cabrera 2017).

Grandmothers as Human Alloparents
If you compare the senescence of physiological functions in human females, you see a remarkable pattern.
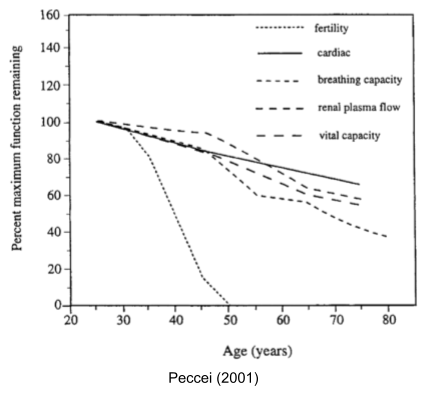
For the vast majority of animals, death occurs very shortly after infertility. Infertile organisms live in the selection shadow, where somatic maintenance is deprioritized. And early menopause is exceedingly rare in the animal kingdom; only found in a handful of other species.
In preindustrial high infant mortality regimes, grandmothers consistently have a tremendous influence on the reproductive success of their offspring. Post-reproductive women gained roughly two extra grandchildren for every ten years they survived past completion of their childbearing (Hrdy 2009; Sear & Mace 2008). Ethnographers report grandmothers are economically productive, provide childcare, and also play a role in teaching motherhood skills. These data suggest that early menopause evolved in the human transition to cooperative breeding.
What conditions would cause a great ape to evolve (grandmother-based) cooperative breeding? We might expect three:
- Environmental scarcity. Cooperative breeding tends to emerge when mothers cannot raise children alone. This is consistent with the Kingdon (2000) hypothesis that Erect speciation occurred in a desert (e.g., Near East), an argument grounded in our hairlessness and moisture-retaining nose,
- Reduction in infanticide rates. Hyper-possessive behaviors are a maternal strategy of last resort. Offspring have more latitude when the risk of male infanticide is low. Whatever the reason, humans exhibit low infanticide rates. This development almost certainly predates and prefigures cooperative breeding.
- Closeness of maternal kin. One of the central objections to the cooperative breeding hypothesis is that both great apes and hunter-gatherers appear to be patrilocal (male bonded). How could selection for grandmothering occur if females disperse away from their own kin? One could perhaps appeal to sororal polygyny as a stepping stone. But Alvarez (2004) showed that most hunter-gatherer groups are actually bilocal, not patrilocal. Perhaps female dispersal was sufficiently flexible to allow female kin.
An Avian Pathway
What drives species towards these archetypes? We don’t have a complete theory. But both multilevel societies (MLS) and cooperative breeding (CB) tend to require environmental harshness, and kinship-bonded, socially tight knit groups.
Across all mammalian species, less than 1% exhibit CB strategies. A similar fraction of species manifest as MLS. And despite their overlapping drivers, finding species with both archetypes is very unusual in mammals (but see Ren 2012; Xiang et al 2019).
So how did humans evolve both?
Birds may provide some insight. In them, cooperative breeding occurs more frequently (~8%). And MLSs and CB frequently co-occur in birds (Camerlenghi et al 2021).
Computational modeling suggests that these high sex ratios can incentivize a behavior change away from multiple-mating and towards pair-bonding (Coxworth et al 2015). If hominin evolution followed this sequence, this would suggest that early menopause induced our species to enter the bonding pathway.

What to expect from an animal who took the Avian Pathway?
Suppose you construct a list of anatomy, behaviors, and cognitive affordances of humans. Then make the same list for chimpanzees, and categorize each element as shared.
- Shared phenotypes. Both species engage in mindreading, toolmaking, sophisticated coalitionary politics, engage in warlike raids, rudimentary culture…
- Derived phenotypes. Only humans have the capacity for language, prestige status psychology, altruism, explosive cumulative culture…
The list of shared phenotypes is a useful reminder of our lowly origin. But the list of idiosyncratically-human phenotypes (aka human universals). is remarkably long. Chimpanzees and humans share a common ancestor some 7-9 mya, a considerable amount of time. But why do humans have these derived features, and not others?
While the MLS-CB social phenotype doesn’t explain the origin of all human universals, it does make our phenotype much less surprising. Let me illustrate this in the domains of language, prestige, and cumulative cultural evolution.
As many theorists have noted, language could not evolve without shared intentionality (the capacity to share attention on a single option), altruism (information donation is an altruistic act), and vocal control (to support e.g., babbling). None of these capacities exist in great apes. But these prerequisites are likely to exist in a cooperative breeding species!
Most primates grapple for status using dominance psychology, which involves aggression-fueled contests. But humans also gain status by prestige, a status system grounded in admiration and deference. We can see these two rival systems expressed in politics. Big men societies use prestige-based leadership; chiefdoms use dominance-based leadership. Both can be seen in eye gaze behavior. In a dominance context, eye contact is confrontational; in prestige contexts this same behavior is deferential.
Henrich & Gil-White (2001) view prestige as a uniquely-human psychological adaptation to facilitate cultural transmission in humans. But one other species seems to have evolved a form of prestige as well: the Arabian babbler. It competes for the right to perform altruistic acts, such as feeding each other, territory defense, and sentinel guard duty (Dattner et al 2015; but see Wright et al 2001). Zahavi describes this prestige behavior using the handicap principle: only truly fit individuals can afford to produce these costly signals of altruism.
Perhaps prestige psychology will soon be found in other cooperative breeding species too. Or perhaps prestige evolves in response to unusually high cost (often mortal) dominance contests, which dovetails nicely with the notion that human reverse dominance psychology evolved in response to projectile weapons.
Less is known about the cognitive correlates of multilevel social organization. A research area to watch closely.
Humans do have an unusually robust capacity for cumulative cultural evolution (CCE); this trait is often credited for our ecological dominance. Undoubtedly, language supercharged our cultural capacities. The population size hypothesis suggests that demographic factors also constrain CCE (Derex & Mesoudi 2020). Multi-level societies have much larger effective group sizes, and group size is an accelerant. Further, the social network architecture inherent in MLS societies can also facilitate cultural recombination (Cantor et al 2021). Taken together, the human shift towards MLS organization seems to have prefigured our later cultural superpowers.
Until next time.
References
- Alvarez (2004). Residence groups among hunter-gatherers: a view of the claims and evidence for patrilocal bands
- Burkart & van Schaik (2010). Cognitive consequences of cooperative breeding in primates?
- Cantor et al (2021). Social network architecture and the tempo of cumulative cultural evolution
- Camerlenghi et al (2021). Cooperative breeding and the emergence of multilevel societies in birds
- Chapais (2008). Primeval Kinship
- Coxworth et al (2015). Grandmothering life histories and human pair bonding
- Dattner et al (2015). Competition over guarding in the Arabian babbler (Turdoides squamiceps), a cooperative breeder
- Derex & Mesoudi (2020). Cumulative cultural evolution within evolving population structures
- Henrich & Gil-White (2001). The evolution of prestige: freely conferred deference as a mechanism for enhancing the benefits of cultural transmission.
- Heinrich (2012). The puzzle of monogamous marriage
- Heinrich (2020). The WEIRDest People in the World: How the West Became Psychologically Peculiar and Particularly Prosperous
- Hrdy (2009). Mothers and Others
- Hrdy & Burkart (2020). The emergence of emotionally modern humans: implications for language and learning
- Kingdon (2000). Lowly Origin: where, when, and why our ancestors first stood up
- Marlowe (2003). The Mating System of Foragers in the Standard Cross-Cultural Sample
- Sear & Mace (2008). Who keeps children alive? A review of the effects of kin on child survival.
- Sherman et al (1995). The Eusociality Continuum
- Peccei (2001). Menopause: adaptation or epiphenomenon?
- Ren (2012). Evidence of Allomaternal Nursing across One-Male Units in the Yunnan Snub-Nosed Monkey (Rhinopithecus Bieti)
- Tomasello & Gonzalez-Cabrera (2017). The Role of Ontogeny in the Evolution of Human Cooperation
- Qi et al (2017). Male cooperation for breeding opportunities
- Wilson (1975). Sociobiology: the new synthesis
- Wright et al (2001). Cooperative sentinel behaviour in the Arabian babbler
- Xiang et al (2019). Routine allomaternal nursing in a free-ranging Old World monkey
- Zahavi (1995) Arabian babblers: the quest for social status in a cooperative breeder
