
Part Of: Anthropogeny sequence
Content Summary: 6000 words, 30 min read
What is a Multilevel Society?
Many mammals live with others. But for the vast majority of these species, social life revolves around families, or groups – not both.
In group-living species, there are no stable breeding bonds. In such multi-male, multi-female (mm-mf) groups, mating is promiscuous. For example, Wrangham estimates that female chimpanzees copulate between 400 and 3,000 times per conception, and female bonobos between 1,800 and 12,100 times. In such species, children bond with their mother, but cannot hope to recognize their father (who could be any of the males within the group).
In family-living species, the family lives autonomously. There are two prominent kinds of primate family: pair-living families (e.g., gibbons), or in one-male units (OMUs) with one male and several females.
But some thirteen primate species (5%) live in multifamily groups. For these species, families (polygynous OMUs) not only share the same space, but also participate in group-level relationships and behaviors.
While some of these multifamily species exhibit two levels (family and clan), other species have more. Sometimes clans coalesce as multilevel societies – apex levels defined first as spatial tolerance, then full-fledged social affiliation.
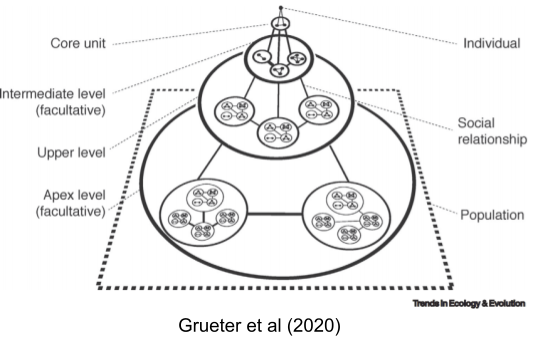
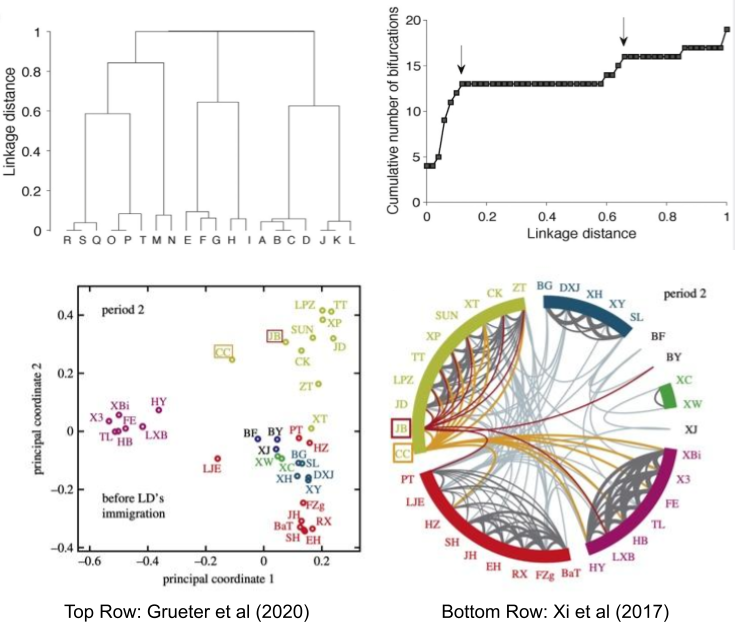
These concepts have been operationalized. By GPS tagging individual primates, proximity data can empirically demonstrate the existence of such levels:
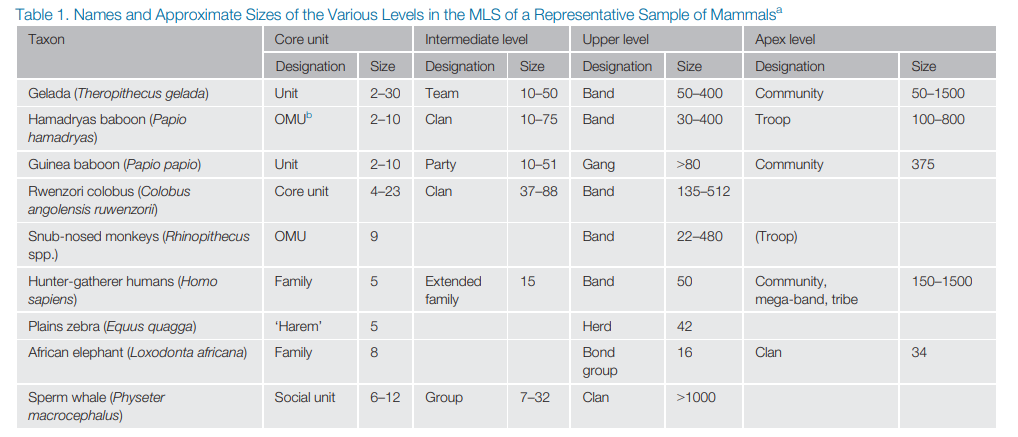
Only a few species live as multi-family or multi-level societies. Here is a partial list from Grueter et al (2020):
These species come from a wide diversity of taxa, suggesting that multi-level sociality is a derived trait
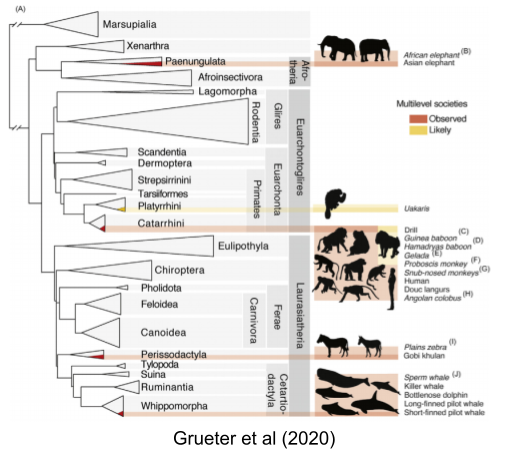
Humans are one example of MLS primate. Today, we’ll dive into nonhuman primates. By understanding these species on their own terms, we should improve our ability to understand not just how mammalian societies function, but also gain insight into how our own species evolved.
So let’s dive in! Our exemplars come from two subfamilies within the Old World Monkeys:
Cercopithecines:
- Geladas (Theropithecus gelada)
- Hamadryas baboons (Papio hamadryas),
- Guinea baboons (Papio papio), discussed occasionally
Colobines:
- Golden snub-nosed monkeys (Rhinopithecus roxellana).
- Rwenzori colobus (Colobus angolensis ruwenzorii), discussed occasionally
- Proboscis monkeys (Nasalis larvatus), discussed occasionally
MLS Social Organization
Kappeler & van Schaik (2001) note that primate social systems rest on three pillars:
- social organization (typical group size, composition, spacing, and dispersal patterns of a given species)
- mating system (patterns of sexual behavior)
- social structure (patterns of relationships between individuals)
Let’s discuss social organization first.
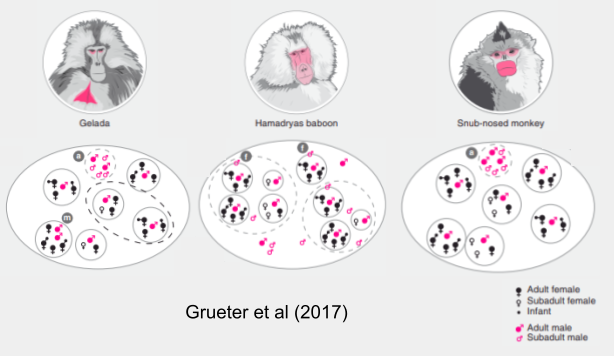
- Hamadryas baboons have all four layers: OMUs, clans, bands, and troops. These layers generate a multilevel allegiance system which mirrors the complexities of a human tribe. Hamadryas clans in the same band mingle while foraging, but males ally with their own clan members in a fight. Members of different clans in the same band will in turn unite against members of alien bands.
- Geladas only have two stable layers: OMUs and bands. Rarely, when an OMU experiences binary fission, the two separated units may cooperate to form a team (Snyder-Mackler et al 2011), by virtue of the bonds of between-OMU female kinship. Occasionally, geladas bands come together, at least spatially, into apex level communities.
- Golden snub-nosed monkeys have three layers. Many OMUs consistently congregate as bands. Every winter, when local food density peaks, these bands fuse into a single troop (Qi et al 2014).
Anthropoid primates typically features unisexual dispersal: one sex disperses to preclude inbreeding, the other remains with its natal group (is philopatric). The latter typically has a kinship advantage: it maintains lifelong social ties with their same-sex relatives.
In several MLS societies, bisexual dispersal occurs – in general, this dispersal regime tends to correspond with strong male-female bonds. Hamadryas baboons are predominantly male philopatric: most often the females disperse. Gelada baboons are female philopatric, and the males disperse. Snub-nosed monkeys are also female philopatric, and the males disperse.
Polygynous species by definition contain a sizable number of bachelor males. Bachelors have up to three “career paths” available:
- Some bachelors join OMUs as followers. Leaders tolerate followers because they are often kin, and also because follower males defend against takeovers, effectively increasing OMU longevity. Followers don’t immediately receive mating access, but they do receive other benefits, detailed below.
- In male philopatric MLS species (e.g., hamadryas and Guinea baboons), non-follower bachelors typically become solitary.
- In female philopatric species (e.g., geladas, snub-nosed monkeys), bachelors can choose a solitary life, but may instead join an all-male band (AMU), which poses an increased threat to OMU security.
In sum, here is how these social organizations differ (more contingent higher levels patterns are omitted for simplicity):
Novel Social Signatures
What are the cognitive demands for life in a multilevel society? For this, we begin with a primate whose cognition we understand quite well: Homo Sapiens.
Human friendships range from casual friends to more intimate associations. But the amount of time we invest in relationships is not continuously graded. Instead, ego networks naturally cluster into four separate groups: support clique (~5 people), sympathy group (~15 people), affinity group (~50 people) and affinity network (~150 people). Willingness to act altruistically tends to stop here, but the number of acquaintances people typically have is ~500, and the number of faces we can recognize may be bounded at ~1500.
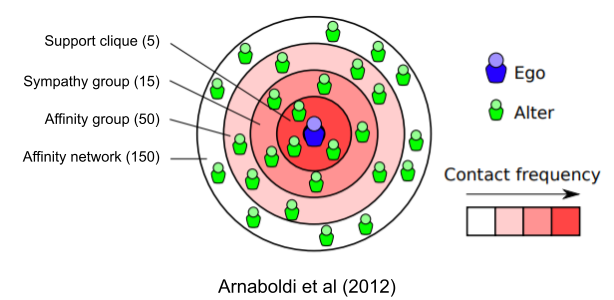
These Dunbar graphs also manifest as typical sizes of human groups (Dunbar 2020); indeed, groups that don’t conform to these sizes deteriorate more quickly (Dunbar & Sosis 2018). Human groups have four nested components (family, group, clan, tribe) – is it really so surprising that we have four kinds of friends (best friends, close friends, friends, acquaintances)?
A social signature is the distribution of social effort that people invest in their friends. Individual social signatures are resistant to turnover: the distribution of our social effort doesn’t change even when we lose or add new friends (Saramaki et al 2014). But the modal social signature for multilevel primates converges on four clusters.
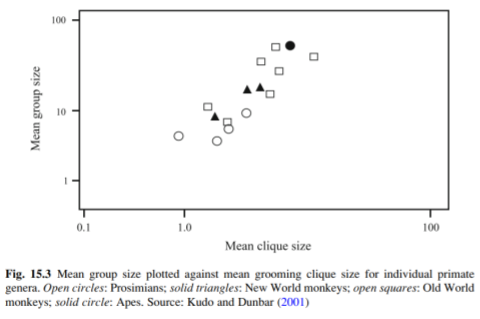
What about the social signature for other primates? Kudo & Dunbar (2001) note
For the majority of the species in this sample, the mean size of networks (i.e. the number of animals linked together by a continuous chain of relationships at the defined discriminant level) is typically around 75% of total group size. This suggests that the majority of individuals are linked together in a single network, with a small number of attached peripheral individuals. These individuals either lead a solitary existence within the group or are members of very small peripheral networks.
Within these bonded networks, primates do develop unusually strong ties to a handful of conspecifics. These cliques are analogous in size and quality to best friendships we see in human beings:
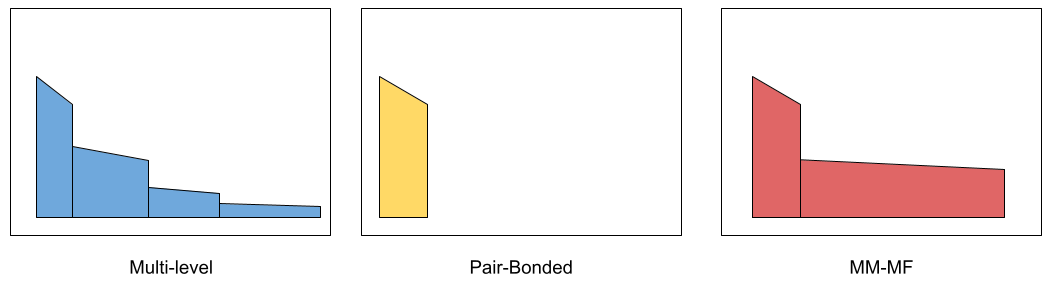
This data (and the agent-based modeling of Sutcliffe et al 2016) suggests three signature archetypes across the primate order:
One benefit from the MLS acquaintance layer is plausibly information transfer. In humans this hypothesis has been explored in seminal Granovetter (1973) The Strength of Weak Ties. Henrich’s excerpt Collapse of Supermind attests to the significance of social network size to sustain cumulative culture.
MLS Mating System
On to the second pillar: mating systems!
For bachelors, there are up to four routes to reproductive success: the initial unit strategy, where a follower befriends (or solitary kidnaps) a juvenile female. The takeover strategy involves physical combat. Followers have access to two other strategies: the opportunistic strategy where you gain a harem when the leader male isn’t around, and the inheritance strategy where females are peacefully transferred from leader to follower. To illustrate how takeovers often go, here’s Pines et al (2011) describing an April 2008 hamadryas takeover.
Takeover of ‘‘Lizzy’’ (adult female with older infant) from ‘‘Pete’’ (old leader) by ‘‘Skivy’’ (subadult solitary): Skivy was observed following Pete and his female Lizzy (Pete’s only remaining adult female). Flanked by ‘‘Herb’’ (a deposed leader with similar facial features to Pete who became Pete’s follower after his own loss of females described below) and by ‘‘Feet’’ (a young subadult follower of Pete), Pete and Lizzy were observed hastily fleeing from Skivy, who followed but did not physically interact with Pete, Herb, or Feet. Skivy continued to pursue Pete, Lizzy, and Feet the next day. On one occasion that Skivy got close to Pete and his entourage, Feet turned and chased Skivy. Approximately half an hour after this, and with Feet no longer around, Skivy approached the foraging Lizzy and grabbed her. Pete, who was about 8 m away, picked up Lizzy’s infant and fled. After a brief flurry of mounts and grooming, Skivy repeatedly herded Lizzy toward Pete and continued to mount and groom her in full view of the now deposed leader.
From a comparative perspective, hamadryas baboons have unusually enhanced levels of sexual coercion. They exhibit herding: keeping their females within 2m using e.g., neck-biting behaviors (Swedell & Schreier 2009). Male aggression peaks immediately after a takeover, and may function to condition a female. Other species have more robust expressions of female choice. Golden snub-nosed monkeys use paternity confusion to preclude takeover-driven infanticide. Guinea baboon females don’t experience “takeovers” at all; but simply move to different OMUs if their current male doesn’t suit them.
In contrast with most mm-mf species, males in multi-level societies associate closely with females at all times. This contrasts with mm-mf primates, in which males consort and mate guard only when females are in estrus.
MLS mating systems varies along other dimensions as well:

Intense Sexual Selection
Sexual selection can be usefully decomposed into intrasexual selection (male-male, female-female), and intersexual selection (male-female). From a male-male competition perspective, strategies vary according to social structure:
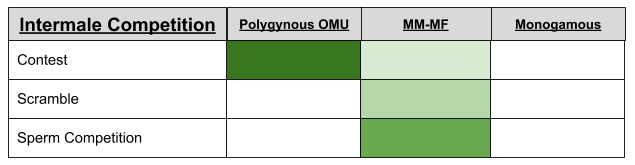
In all primates, testes size correlates with body size. However, after controlling for body size, mm-mf species have larger testes in men. This is because females have multiple copulations during estrous, and the male who delivers the most gametes has a fitness advantage (hence the arms race to produce more gametes). For example, despite being much smaller than humans, chimp testes are much larger.
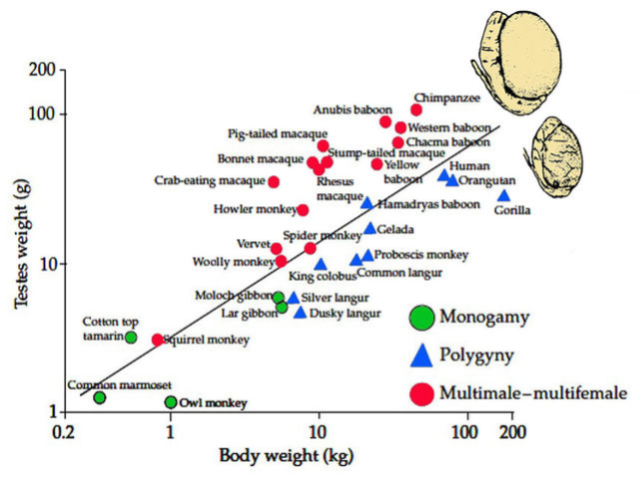
In polygynous societies, where a single male forms stable breeding bonds with multiple females, males don’t have to worry about competing with other males’ sperm. But they do have to worry about bachelors challenging & overtaking their harem. And as harem sizes grow larger (bigger operational sex ratio OSR), bachelor threat becomes increasingly intense (this is polygyny’s math problem). Size and weaponry drive success in contests for sexual access. As the intensity of bachelor contests increases, we see more sexual dimorphism: males whose bodies (and teeth!) are bigger than women (Clutton-Brock et al 1977).
That’s how male-male sexual selection works in traditional primate societies. But what happens in a multilevel sociality? More specifically, what happens when you take spatially autonomous families, and have them co-reside. Two effects manifest immediately:
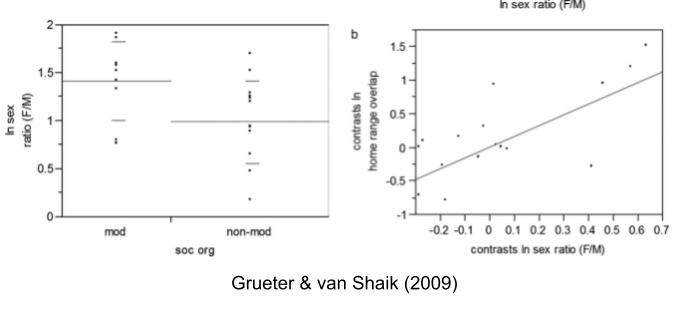
- Nearby OMU males often exacerbate opportunities for contest competition. And Grueter & van Schaik (2009) found that, indeed, sexual dimorphism is even greater in multilevel primates than traditional polygynous societies.
- Nearby OMU males often provide females with more opportunities for extra-pair mating (EPM). In polygynous societies, male-driven infanticide is a constant threat (killing an infant terminates lactation, and thereby accelerates oestrus). “Cheating” promotes paternity confusion, and reduces the infanticide threat. Qi et al (2020) found that, while no observed instances of EPMs occurred, paternity tests revealed more than half of all children were sired by non-resident males.
But not just the OMU males that change the sexual selection calculus. Bachelors behave differently in multilevel societies as well! Due to the unusually tolerant male-male dispositions required in multilevel societies, and the dramatic expansion of male kin groups outlined above, bachelor males live in unusually cooperative all-male units (AMUs). OMUs congregate together in breeding bands (BBs), AMUs congregate in all-male bands (AMBs).
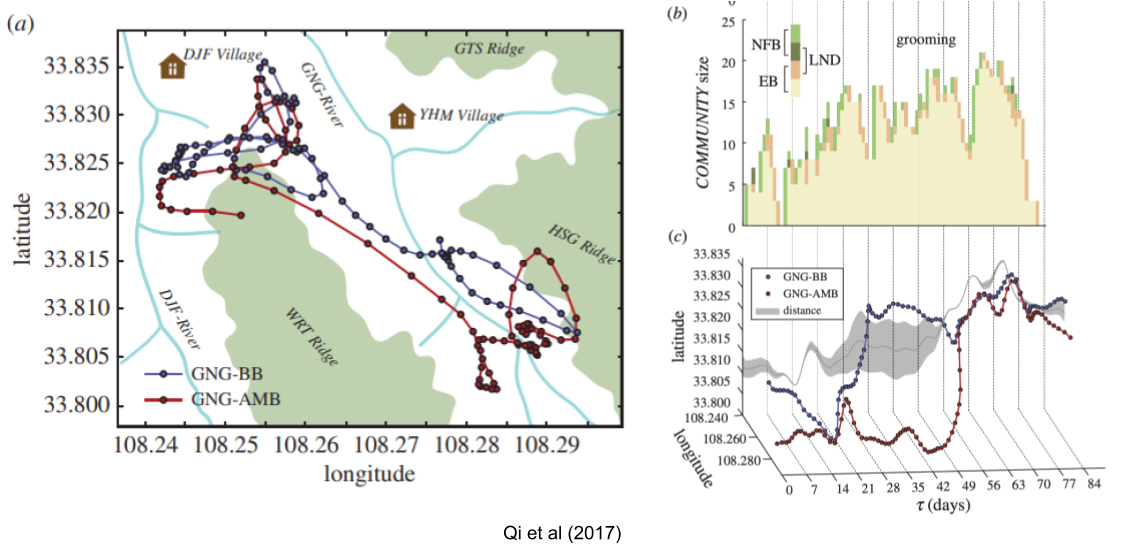
For colobus monkeys, Qi et al (2017) showed that AMBs tend to be organized along kinship lines. Their movements tend to shadow that of the breeding band. What’s more, social patterns of grooming tend to correlate with distance between these two groups – this is likely associated with preparations for violence.
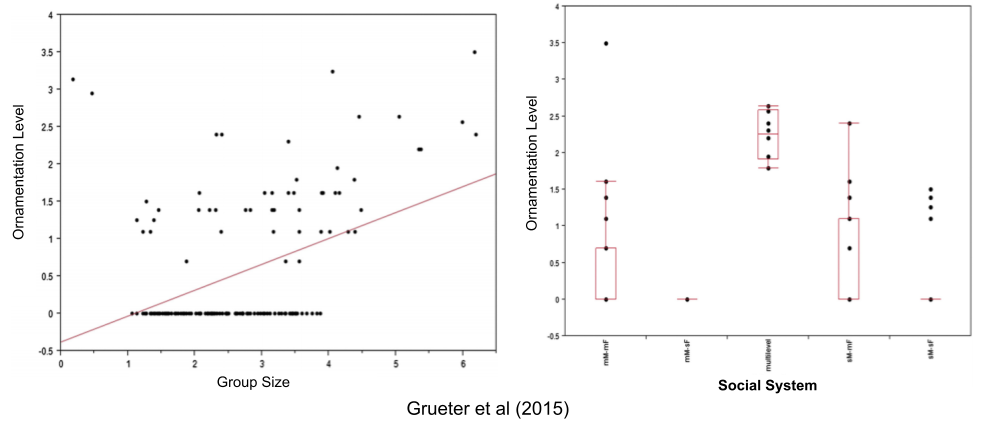
Finally, let’s consider secondary sexual traits (ornaments). These come in three varieties: hairy traits (capes, tufts, beards), fleshy traits (lips, nose, humps), and colorful traits (red hair, blue scrotum, etc).

Such ornaments serve at least one of the following functions
- Status Signaling. Some ornaments signal dominance between males.
- Attractiveness. Some male ornaments are associated with female choice.
In traditional social structures, individuals are well-known to one another. In multilevel societies, which occasionally coalesce into very large groups, anonymity is more prevalent. In contexts of limited social knowledge, the functional value of such ornaments is particularly high. And indeed, multilevel societies are disproportionately likely to exhibit these ornaments.
MLS Social Structure
We’ve discussed social structure and mating systems. Time for the third pillar: social structure.
In hamadryas societies, if a leader male is incapacitated, harem females will often go their own way. But in geladas, females in an OMU will stay together even if the male is deposed. This suggests that in gelada OMUs are bonded by female kin; whereas the pair-bond is the primary core unit glue in hamadryas.
You can actually predict this pattern with social network analysis (SNA) to evaluate the social bonds of a given society. Matsuda et al (2012) found that females are more network-central to the colobines, whereas males are more central in cercopithecines. Clustering analysis, however, reveals that geladas OMUs become more unstable when females, but not males, are removed from social networks.
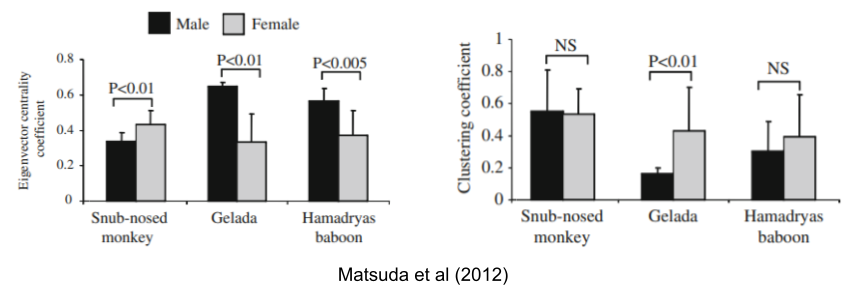
Why should these species differ so radically in social structure? These differences are largely driven by philopatry-based kindreds, sexual selection, and dispersal patterns. Recall that in multilevel societies, dispersal risk is lessened by transferring within the super-group. But which level do individuals transfer? Here’s some data:
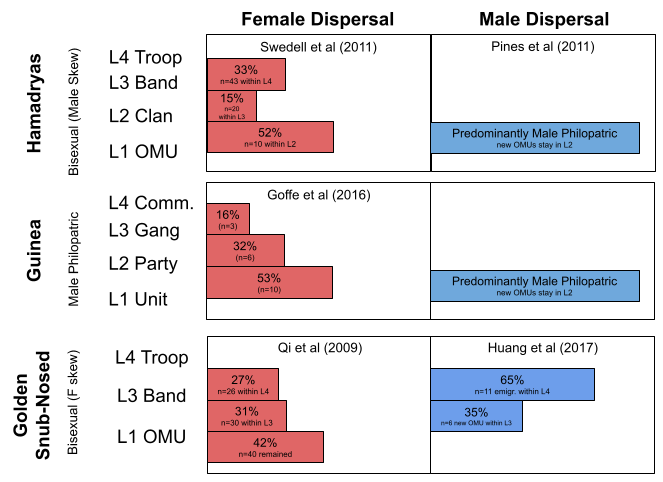
Interestingly, Rwenzori colobines have a pattern of upper-level female dispersal and lower-level male dispersal. This pattern is rather unusual, and closely resembles one other MLS primate species: Homo Sapiens.
These dispersal patterns help explain social structure:
- In hamadryas, male dispersal patterns contribute to the L2 male alliances, used to repulse bachelor males. In this species, male sexual coercion may explain the weak female bonds, even within-harem.
- In geladas, the lack of female transfer within bands and a lack of clan-based male bonds are likely reasons why gelada bands are not maintained as coherently as hamadryas bands.
- In golden snub-nosed monkeys, female philopatry and the colobine penchant for allomothering generates very strong female cohesion within core units (but male-female relationships within OMUs are also quite strong).
Primate societies typically spend their waking days together. But a minority of species (incl. chimps, bonobos and humans) operate under fission-fusion dynamics: subgroups with variable composition forage independently. Fission-fusion behavior is an adaptive response to variable foraging environments, allowing species to dynamically alter their social structure in spatiotemporally variable ecologies. The question of whether fission-fusion behavior requires additional cognitive abilities is an area of live research.
Historically, there has been a strong tendency to conflate fission-fusion with multi-level societies. But there is a key difference: in mm-mf primates that use fission-fusion dynamics, subgroups are formed on an individual basis (they are atomic communities). In contrast, multilevel, fission-fusion primates form subgroups on a familial basis (they are molecular communities). Families are never separated: they forage together. An interesting exception to this trend is human beings, which use an atomic style during the day (by the sexual division of labor, our families aren’t literally inseparable), but a molecular style at night (unlike chimps, human nature has us rather consistently sleeping with our families).
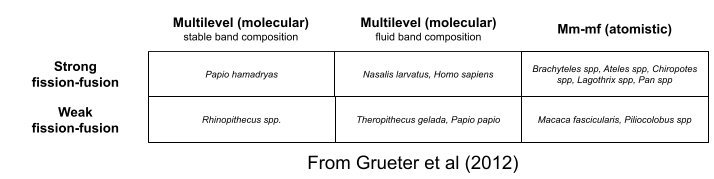
Stronger Kindreds
In comparison to mm-mf groups, the potential for kin selection-based cooperation is dramatically enhanced by multilevel social organization. Let’s dive in.
For nearly all primates, the most common philopatry pattern is female philopatry. Only a few primate species, including our two closest relatives chimpanzees and bonobos, exhibit male philopatry.
Primates are capable of kin recognition. We find evidence for this in nepotism (preferentially helping relatives), incest avoidance (they largely avoid mating with conspecific kin), and dominance relations (vengeance is often directed at the aggressor’s kin). Kin recognition is grounded in parental attachment, and from there an inductive ability to notice adjacent attachment relationships. But kin recognition varies by social structure. Consider the following:
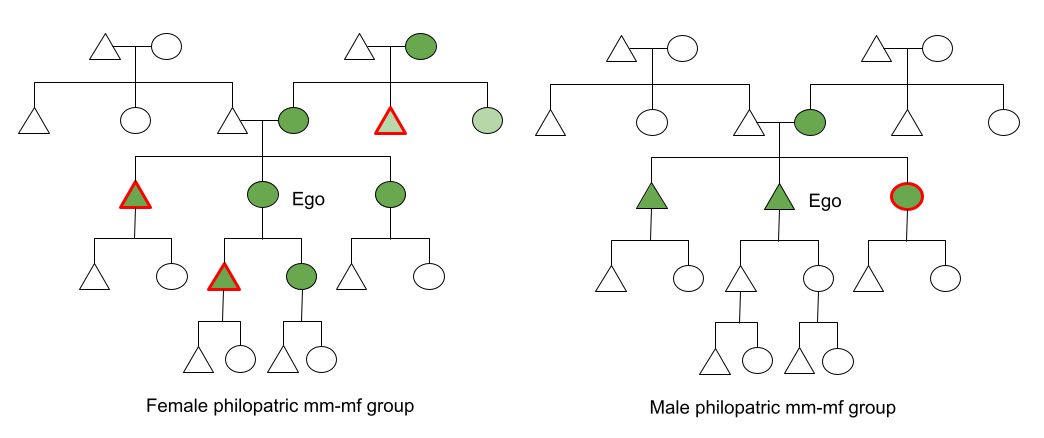
Symbol Key:
- Circle = female; Triangle = male
- Green = fully recognized kin, Light green = imperfectly recognized kin, White = unrecognized kin
- Red Outline = emigrant, Blue Outline = immigrant
In female philopatric mm-mf groups, (e.g., macaques) Ego recognizes her mother and children, by virtue of parturition and lactation. Ego is also able to recognize her siblings (“the other juveniles bonded to my mother”) and maternal grandmother (“the older female bonded with my mother). She may also be able to recognize her maternal aunts and uncles, and sister’s offspring, but this requires a second inferential link, and the evidence for this ability is mixed. Ego’s father does not recognize her (impossible to say who inseminated the mother), and Ego will never know her son or’s brother’s offspring (since they dispersed as juveniles). In fact, the three males she can recognize as kin leave. The female kin she can recognize often cooperate jockeying for dominance status. For all individuals then, matrilines (n=5) are important determinants of social structure.
In male philopatric mm-mf groups (e.g., chimps), the situation is more lonely. Resident male Ego cannot recognize his father, but he also cannot recognize his mother’s kin, because mom is an immigrant. He can recognize his siblings. But he cannot recognize his brother’s kids (even his brothers don’t know). Nor can he build alliances with his sister’s sons, because his sister emigrates. Patrilines (n=2) permeate the society, but given their shallowness, the situation is more akin to “every male for himself”.
The situation changes when pair-bonding is introduced (the advent of multifamily groups).
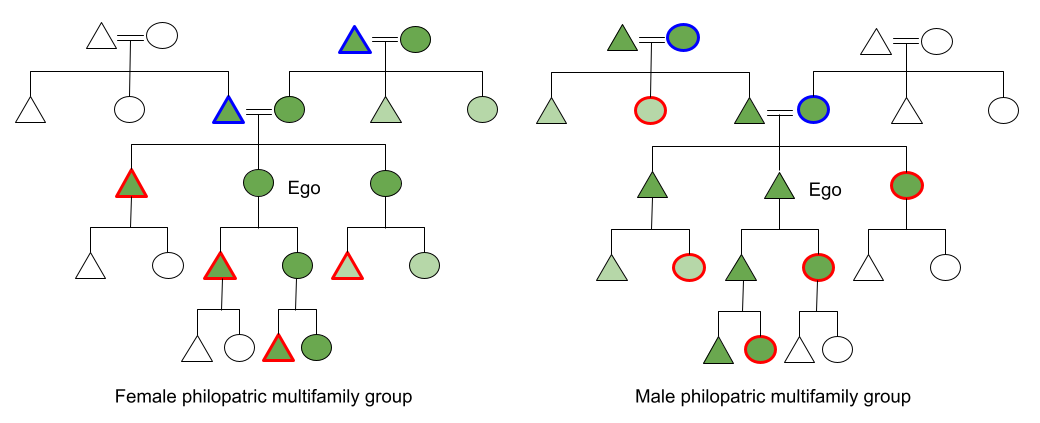
For female philopatric multifamily groups, in addition to the above, Ego gains the ability to recognize & bond with her father and maternal grandfather. These individuals may prove useful allies in her life. But these eureka moments stop their: their immigrant status prevents Ego from meeting this part of her extended family.
For male philopatric multifamily groups (e.g., ancestral humans), we see that pair-bonding has radically improved Ego’s situation. Ego now recognizes his father, and his paternal grandfather (that older male with whom his father is most strongly bonded with). Ego can also bond with, and promote the welfare of, his children, and his (non-emigrated) extended family. He also may form weaker bonds with his brother’s offspring, and paternal uncles & aunts. The scope of patrilines radically expands, rather than a single male ally, Ego’s patriline network has achieved n=6.
In contrast with mm-mf groups, pair-bonding dramatically expands male kin recognition. These kindreds dramatically expand the cooperative potential of agnatic kin.
Two Evolutionary Pathways
Shultz et al (2011) model the evolution of social life within the entire primate order. The earliest primates lived solitary, nocturnal lives. The shift towards diurnal social living represents a major shift in the primate adaptive landscape, with an increased emphasis on visual processing (efficient foraging) and group living (predation mitigation).
Dunbar & Shultz (2010) argue that sociality comes in two forms. Aggregations (siimple, fluid groups) form when individuals benefit from home range overlap. Congregations (complex, bonded groups) require a repurposing of mother-offspring attachment towards others. Such animals make friends (sensu Silk 2002). Most social mammals (e.g., ungulates) live in aggregations. A few mammals (including social carnivores, dolphins, elephants, and anthropoid primates) live in congregations.
For primates, the transition from solitary living seems to have two stages. First, there was a stage with unbonded mm-mf aggregations (eg. diurnal lemurs). This was succeeded by bonded mm-mf congregations.
As mentioned, this escape from solitary lifestyle was likely stabilized by exaptations of mother-offspring attachment. These bonds were further reinforced by a transition from bisexual dispersal to unisexual (typically male) dispersal, which enabled strong (female) kindreds. The fact that there are zero attestations of reversion back to soli, suggesting that the stabilizing role of these mechanisms.
Female philopatry is expressed in OMUs when males successfully monopolize access to females, and mm-mf when they do not. Transitions between these forms may be ecologically mediated; namely, reductions in food density that cannot support large female aggregations. This hypothesis finds support in Barton (1999)’s observation that savanna baboons, which typically form large mm-mf groups, may sometimes subdivide into polygynous groups in harsher conditions.

Shultz et al (2011) document a transition from unstable mm-mf groups to pair living and social monogamy (not to be confused with sexual monogamy). Social monogamy evolves prior to paternal provisioning, and represents an end state. Opie et al (2013) claims that all appearances of social monogamy take place in unstable mm-mf species, all with heightened levels of infanticide risk. In contrast, Lukas & Clutton-Brock (2013) claim that all instances of pair-bonding evolved from solitary species, for the ecological reason of sparse resource → female territoriality → incentivizing male pair-living strategies.
These two accounts have not yet been reconciled. But Kappeler (2014) suggests both pathways might be possible: lemurs show evidence of both group-living and solitary societies; moreover, the resultant monogamous systems have systematically different properties.
What about multilevel societies? How did they evolve?
Consider again our exemplar organisms (plus lesser-known, suspected, and intermediate MLS species located in Pygathris, Nasalis, Madrillus, and Macaca genus). Yellow denotes OMUs, red as stable MM-MFs, and blue as MLS.
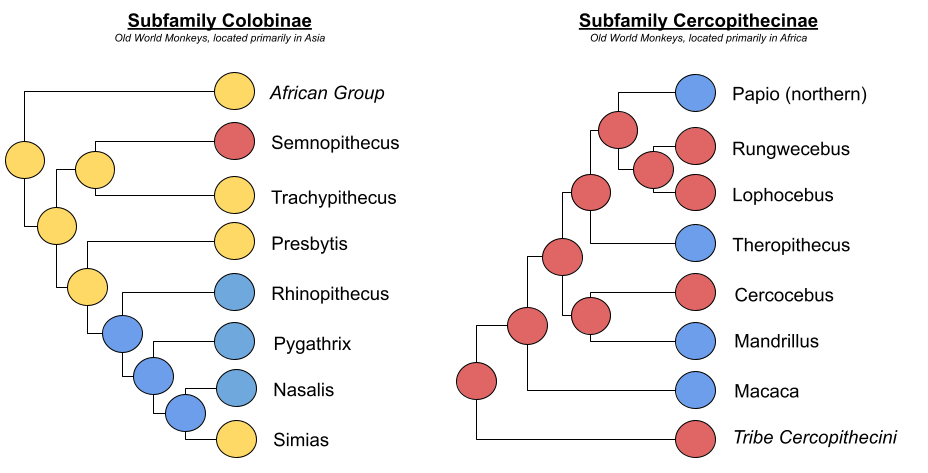
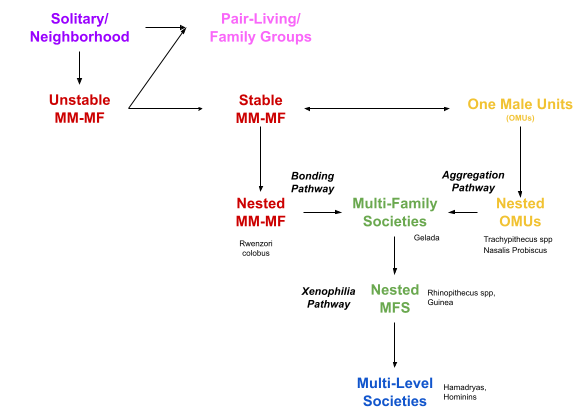
These MLS are derived from different ancestral social systems! Grueter et al (2012) outline two pathways. With the bonding pathway, pair bonding substructures ancestral mm-mf groups. But with the aggregation pathway: autonomous OMUs increasingly overlap, and ultimately affiliate.
Comparative data may allow us to detect substages within these pathways:
First, consider the Rwenzori colobus. This species seems to be a social Archaeopteryx: it represents a transition point halfway along the bonding pathway. The African colobine group contains both OMU and mm-mf species. The Rwenzori is multilevel, but its core units can be either OMU or MM-MF. These core units often “switch modes”. For example, Stead & Teichroeb (2019) document a case where the largest mm-mf core unit, which consisted of 8 adult males and 6 adult females, split into two units: an OMU and a 7-member AMU. It is plausible to speculate that, for the bonding pathway, a multilevel structure predates and incentivizes the ascent of OMUs.
Second, consider the proboscis monkeys. Derived from OMU ancestors, this species appears to be halfway along the aggregation pathway: OMUs are spatially tolerant, but don’t seem socially integrated (nested OMUs). Similar results obtain for several species in the Pythagrix genus.
Third, we’ve already seen how some MLSs have an apex or fourth level (e.g., hamadryas) and some do not (e.g., geladas). This suggests that the apex level affiliation postdates multifamily groups. Let’s call this xenophilia pathway. This pathway too may decompose: from spatially tolerant behavior (nested MFS) towards direct social affiliation (true multi-level societies).
Two Evolutionary Scenarios
What caused these certain colobines and cercopithecines to “take the leap” towards multi-level sociality?
For colobines, an important ecological determinant is diet. Snub-nosed monkeys are unusual in that their diet is dominated by lichens, a low-quality & high-abundance food. For other colobines with different dietary profiles, the amount of scramble competition is high: larger groups means more energy invested in travelling to productive forage areas. But with multilevel colobines like Rhinopithecus bieti, their diet significantly reduces scramble competition (slope is 30x smaller, in this example).
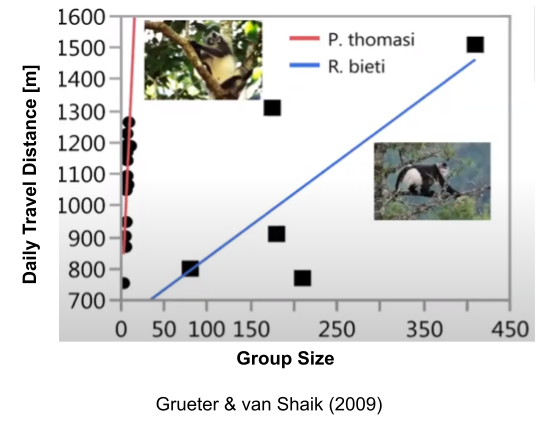
Many other primates share this property of minimal scramble competition, yet they don’t form multilevel societies. It seems that diet is a necessary, but not sufficient precondition.
The bachelor threat hypothesis attempts to explain the snub-nosed monkey phenomenon. Consider the following facts:
- Higher operational sex ratios (OSRs) correlate with increasing OMU overlap (Grueter & van Shaik 2009).
- Multilevel species have higher OSRs (Grueter & van Shaik 2009).
- Infanticide and extra-pair copulation (paternity confusion) are prevalent in these populations.
- Xiang et al (2014) found that colobine males in multilevel societies often collaborate to repulse all-male bands, particularly during mating season.
Perhaps this two-pronged approach explains the evolutionary origins of geladas. Like colobines, geladas feed on low-quality, super-abundant foods (young leaves). This dietary preference dramatically reduces within-group feeding competition. However, geladas don’t suffer as much from bachelor threat. Perhaps this explains why they haven’t taken the xenophilia pathway, in contrast with colobines.
For baboons, Jolly (2020) presents a five-stage reconstruction of Papio evolutionary history, based on modern genetic data.
- Originally two species (Yellow and Kinda) populated South Africa. These baboons lived in a female philopatric, mm-mf configuration.
- As East Africa evergreen forest dried, a pathway opened up. A male-philopatric multi-family ancestral species (gray color, P origin point), mounted a frontier invasion (similar to how starlings colonized North America).
- This multifamily species ultimately diverged into three populations (Guinea, Ancestral Northern, Hamadryas – see Fig C)
- Olive baboons speciate from Ancestral Northern, and revert back to female-philopatric mm-mf, separating Guinea from Hamadryas baboons.
- The species cross-fertilizations in the last step are visible in modern-day “discordant” mitochondrial DNA.
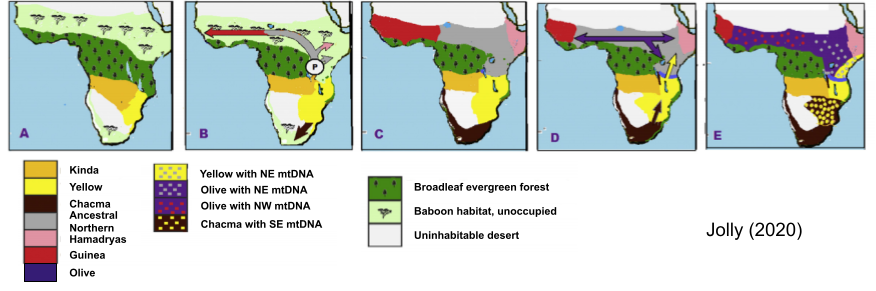
The invasion species P derives from Kinda baboons, which are unusual compared to other baboons for indirect male-male cooperation, as well as stronger male-female relations including comparatively intimate consortships (Petersdorf et al 2019). These features may have produced useful cognitive preadaptations for multi-level social life.
But why would P become male philopatric? Consider a dispersing male living with a group on the frontier. If he disperses away from the (rapidly-moving) frontier, his heritage does not contribute to the subsequent generations of the expanding population. If he disperses into the frontier, he has no group to join, and his chance of starvation or predation is quite high. Finally, if he disperses along the frontier that is also disadvantageous due to the Holt-McPeek effect (Hold & McPeek 1996). Taken together we see that male dispersal is penalized, the dispersal cost-benefit tradeoff changes, and those males predisposed to remain (always a small fraction of a group) would enjoy a fitness advantage. As the frontier expands, typically by group fission seeding the horizon, this process compounds.
Jolly (2020) suggests that this frontier-driven shift to male philopatry allowed these baboons social organization to find a new equilibrium:
Adding male philopatry to preexisting female philopatry would produce troops that were near-endogamous, reducing the effective population size and incurring the risk of inbreeding depression and/or shortage of acceptable mates. At the frontier itself, conditions would minimize this risk. Once the frontier had moved on, however, the male-philopatric troops in the fully occupied landscape behind it would no longer enjoy superabundant resources and would therefore tend to shrink toward a size sustainable as a foraging group within such a setting. Being small and endogamous, however, would imperil a troop’s survival.
This unstable situation changes individual incentives for group encounters:
Troops formed by fusion in this way would maintain their critical size as breeding groups as long as all members frequently gathered at the sleeping site to spend social time communally. At the same time, clans/ parties could forage independently where the ecological setting favored such behavior. In this way, populations that had become male-philopatric remained viable by decoupling the socializing and co-foraging aspects of troop membership
On this hypothesis, the social dispositions of Guinea baboon makes a plausible base from which to derive hamadryas society. In the sparser resources in the Horn of Africa, foraging units would shrink down to the “molecular” OMUs. As the sole defender of their unit, this would have put stress on the males for aggressive protection, and fewer opportunities to interact with individuals outside of their OMU, females would be more socially isolated.
This frontier annealing hypothesis attempts to explain why some baboons embarked on the bonding pathway. But however these northern baboons did it, it’s worth noting the only known case of pathway reversion: olive baboons (Papio anubis), while ancestrally male-philopatric multi-family, reverted back to a male-dispersing mm-mf social organization.
Roots Of Xenophilia
For many primates, xenophobia (between-group hostility) is the norm. For example, as we saw previously, coalitions of chimp males periodically raid neighboring territories, killing anyone unfortunate to cross its path. Xenophobia is largely grounded in resource competition: violence is incentivized if a neighbor possesses a valuable resource, and a group has enough physical power to capture & retain it.
Many intergroup encounters are agonistic. But other encounters are more tolerant. What adaptive value does extra-group affiliation unlock? Examples include:
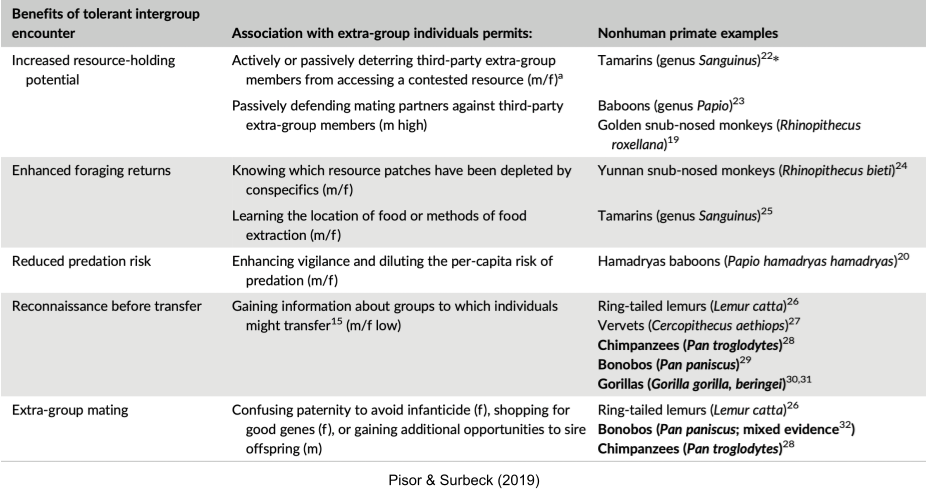
So far, I’ve described the adaptive reasons why group encounters may be hostile vs friendly. But what proximal mechanisms nudge groups in either direction? First, much empirical work has shown relatedness of extra-group members reduces aggression (because of kin selection). Second, familiarity is another key mediator, for at least three reasons:
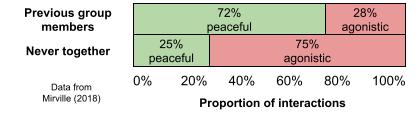
- Post-fission. When a gorilla troop experiences fission, for example, it is much less likely to fight with its previous associates. If the annealing hypothesi is correction, and large group fissions creating MLSs in Papio, this would be how they did it.
- Shared interest. Consider a male philopatric species, where a female leaves Group A and pair-bonds with a male in Group B. Both groups meet from time to time. Such groups have some shared kinship and built-in familiarity via the “linking individual”. Not only this, but conflict that risks harm to the link and her offspring is against the reproductive interests of both the relatives and the affines (“in-laws”). As the number of out-marriages (out-mating + pair-bonding) increase, this reason for cooperation becomes increasingly robust.
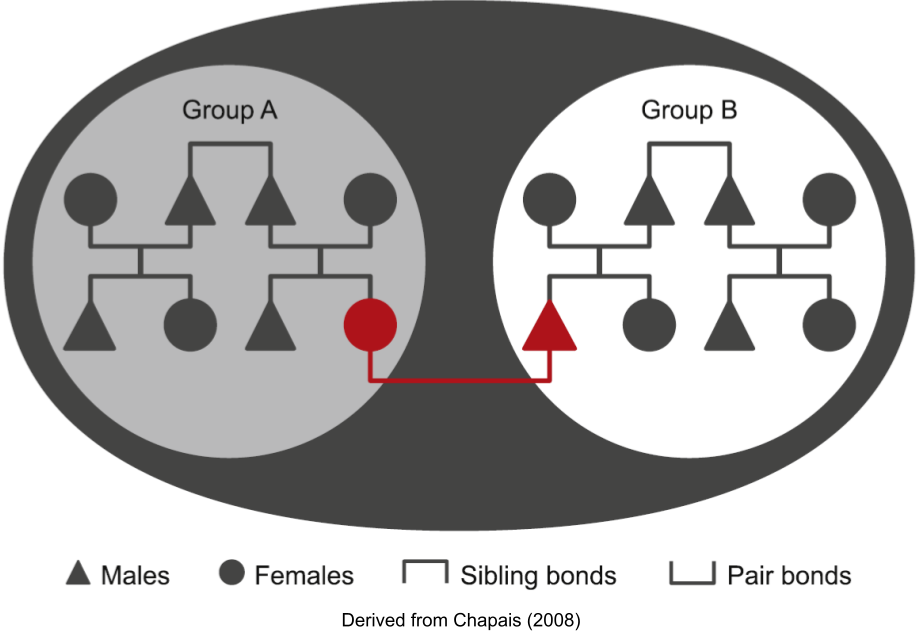
- Mere exposure. Even groups that don’t share many fission- or dispersal-based links may come to act cooperatively, by the brute fact that neutral exposures to strangers paves the way for affinity (in humans, this is known as the propinquity effect).
A summary graphic of our discussion in this section:
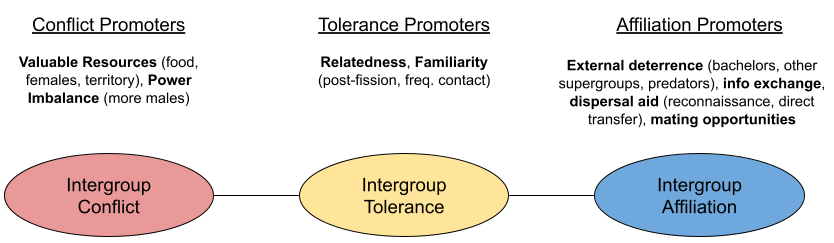
A couple caveats are in order. First, all three affiliation promoters assume frequency of contact: if groups practice avoidance, intergroup relations are moot. Second, the above analysis explores primarily dyadic factors. But the relationship Group B has to Group A may play a role in its receptivity to Group C. Geography plays a role. Some multilevel societies have strictly delineated & coherent apex level structures (e.g., hamadryas baboons); others are more flexible at this higher level (e.g., human beings).
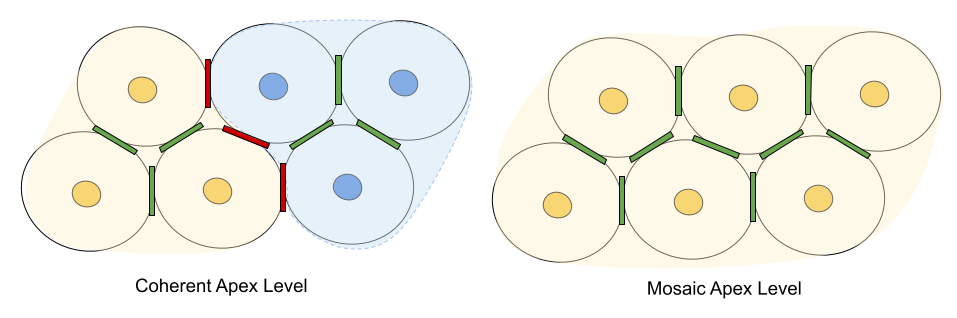
Fry et al (2021) describes peace systems: certain human societies that have managed to avoid war & achieve long-term intergroup peace. And indeed, multilevel societies in other primate societies also seem less prone to intergroup aggression. It seems likely that a deeper understanding of multilevel societies might contribute to furthering the science – and practice! – of peace.
Until next time.
References
Bolded references are ones I found exceptionally interesting.
- Arnaboldi et al (2012). Analysis of Ego Network Structure in Online Social Networks
- Barton (1999). Socioecology of baboons: the interaction of male and female strategies
- Clutton-Brock et al (1977). Sexual Dimorphism, socionomic sex ratio and body weight in primates
- Chapais (2008). Primeval Kinship
- Dixon & Vasey (2012). Beards augment perceptions of men’s age, social status, and aggressiveness, but not attractiveness
- Dunbar & Shultz (2010). Bondedness and sociality
- Dunbar & Sosis (2018). Optimising human community sizes
- Dunbar (2020). Structure and function in human and primate social networks: implications for diffusion, network stability and health
- Fischer et al (2016). Charting the neglected West: The social system of Guinea baboons
- Fry et al (2020). Societies within peace systems avoid war and build positive intergroup relationships
- Goffe et al (2016). Social relationships of female Guinea baboons (Papio papio) in Senegal
- Grannovetter (1973). The strength of weak ties.
- Grueter & van Schaik (2009). Sexual size dimorphism in Asian colobines revisited
- Grueter & van Shaik (2009). Evolutionary determinants of modular societies in colobines
- Grueter et al (2012). Evolution of Multilevel Social Systems in Nonhuman Primates and Humans.
- Grueter et al (2015). Are badges of status adaptive in large complex primate groups?
- Grueter et al (2017). Multilevel societies
- Grueter et al (2020). Multilevel Organisation of Animal Sociality
- Holt & McPeek (1996) Chaotic population dynamics favors the evolution of dispersal.
- Huang et al (2017). Male Dispersal Pattern in Golden Snub-nosed Monkey (Rhinopithecus roxellana) in Qinling Mountains and its Conservation Implication
- Jolly et al (2020). Philopatry at the frontier: A demographically driven scenario for the evolution of multilevel societies in baboons (Papio)
- Kappeler & van Schaik (2001). Evolution of primate social systems
- Kappeler (2014). Lemur behaviour informs the evolution of social monogamy
- Kirkpatrick & Grueter (2010). Snub-nosed Monkeys: Multilevel Societies across varied environments
- Kudo & Dunbar (2001). Neocortex size and social network size in primates
- Layton et al (2012). Antiquity and Social Functions of Multilevel Social Organization Among Human Hunter-Gatherers
- Lukas & Clutton-Brock (2013) The evolution of social monogamy in mammals.
- Matsuda et al (2012). Comparisons of Intraunit Relationships in Nonhuman Primates Living in Multilevel Social Systems
- Miller et al (2014). Diet and Use of Fallback Foods by Rwenzori Black-and-White Colobus (Colobus angolensis ruwenzori) in Rwanda: Implications for Supergroup Formation
- Mirville et al (2018). Low familiarity and similar ‘group strength’ between opponents increase the intensity of intergroup interactions in mountain gorillas (Gorilla beringei beringei)
- Opie, C. et al. (2013) Male infanticide leads to social monogamy in primates.
- Petersdorf et al (2019) Sexual selection in the Kinda baboon
- Pines et al (2011). Alternative Routes to the Leader Male Role in a Multi-Level Society: Follower vs. Solitary Male Strategies and Outcomes in Hamadryas Baboons
- Pisor & Surbeck (2019). The evolution of intergroup tolerance in nonhuman primates and humans
- Qi et al (2014). Satellite telemetry and social modeling offer new insights into the origin of primate multilevel societies
- Qi et al (2017). Male cooperation for breeding opportunities
- Qi et al (2020). Multilevel societies facilitate infanticide avoidance through increased extrapair matings
- Saramaki et al (2014). Persistence of social signatures in human communication
- Shultz et al (2011). Stepwise evolution of stable sociality in primates
- Silk (2002). Using the ‘f’-word in primatology.
- Stead & Teichroeb (2019). A multi-level society comprised of one-male and multi-male core units in an African colobine (Colobus angolensis ruwenzorii)
- Snyder-Mackler et al (2011). Defining Higher Levels in the Multilevel Societies of Geladas (Theropithecus gelada)
- Sutcliffe et al (2016). Modelling the Evolution of Social Structure
- Swedell & Schreier (2009). Male Aggression towards Females in Hamadryas Baboons: Conditioning, Coercion, and Control
- Swedell et al (2011). Female ‘‘Dispersal’’ in Hamadryas Baboons: Transfer Among Social Units in a Multilevel Society
- Xiang et al (2014). Males collectively defend their one‐male units against bachelor males in a multi‐level primate society
