Part Of: Biology sequence
Content Summary: 1400 words, 14 min read.
Ernst Mayr once wrote that “the search for homologous genes is quite futile except in very close relatives”. But evolutionary developmental biology (aka evo devo) has turned this common knowledge on its face. All complex animals – flies and flycatchers, dinosaurs and trilobites, flatworms and humans, share a common genetic toolkit of that govern the formation and patterning of their bodies and body parts.
Let’s dive in.
Principles of Development
Bodies are not built at random. They are constrained by two important organizing principles.
- Bilateral symmetry. The left and right sides of our bodies tend to mirror one another.
- Modularity. Our genomes tend to build recurring segments, and then proceed to customize each segment.
Modularity is one of the most important principles in anatomy. In protostomes like worms and centipedes, modules are expressed in repeating segments. In deuterostomes like penguins and humans, modules are expressed in repeated somites (e.g. vertebrae).

Consider the following facts:
- Trilobite anatomy features many identical legs. In contrast, its descendants (e.g., crayfish) have fewer, and highly specialized appendages.
- Early teeth in e.g., sharks were numerous and undifferentiated. Contrast this with the horse, which has incisors, canines, premolars, and molars.
Williston’s Law generalizes such observations. Earlier species have many, unspecialized modular repetitions. Over time, there is a trend towards fewer, increasingly specialized parts.

Hox Genes and Localization
The genome contains coding genes, which directly encode proteins, and also regulatory genes that modify the activation profile of those coding genes. Regulatory genes form a regulatory hierarchy, whereby gene activation is controlled by increasingly specific activation profiles. The process is an abstraction hierarchy, such as feature hierarchies found in convolutional neural networks. This regulatory system learns how to e.g., deploy calcium proteins in areas where bone formation is prescribed.
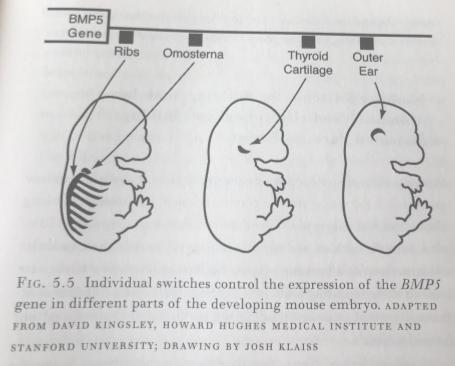
Recall that every cell in an organism contains the exact same DNA. How then does one cell know to become an eye tissue, and another cell knows to become liver tissue? How is cellular differentiation possible?
In order to learn what kind of cell it is, a cell must learn where it is: differentiation requires localization. Roughly speaking, a cell will manufacture eye-specific proteins once it knows that it is located above the nose, and between the ears.
How do cells learn their position? One bit at a time. Per the intension-extension tradeoff, as cells get more location information, their localization window shrinks.
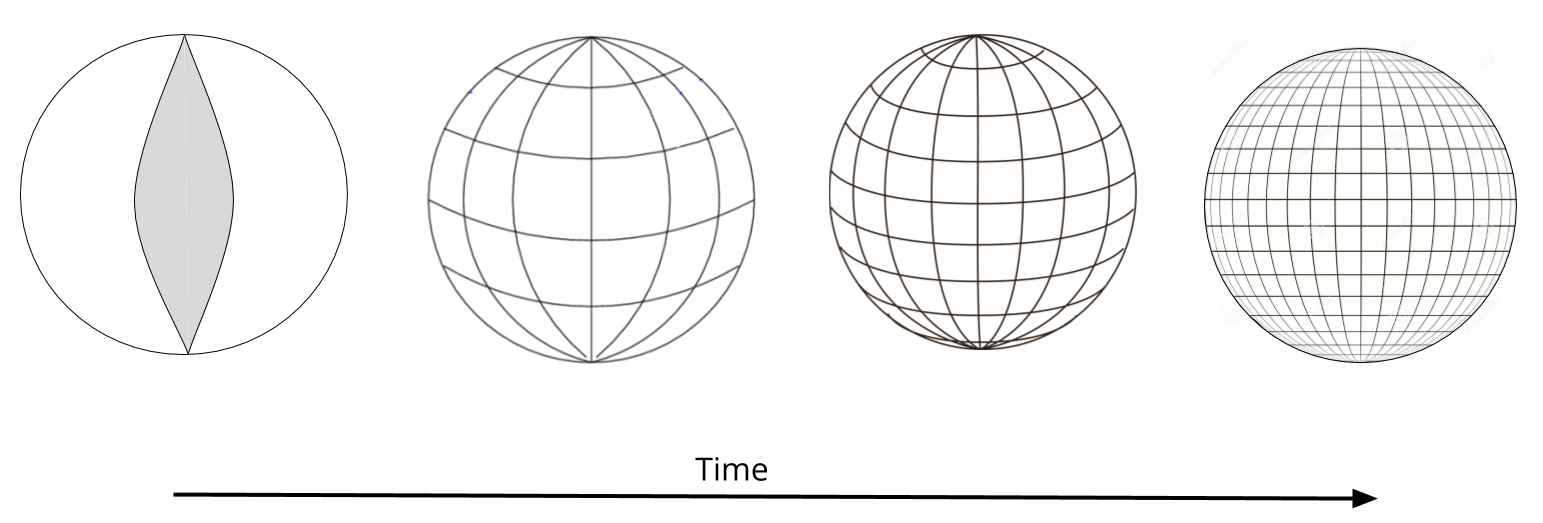
All this is nice in theory, but how does it work in practice?
Just as brain lesions shed light on neuroscience, birth defects shed light on developmental biology. Biologists have been particularly interested in homeotic mutations: mutations that cause body structure to grow in “the wrong place”. Examples include extra fingers in humans, only one central eye in sheep, and legs in the place of eyes in the fruit fly.
A closer look has revealed that homeotic mutations are caused by damage to a specific set of genes: homeobox (Hox) genes. These genes (near the top of the regulatory hierarchy) encode location information, and are conserved across species – the same genes exist in a mouse and a fruit fly:
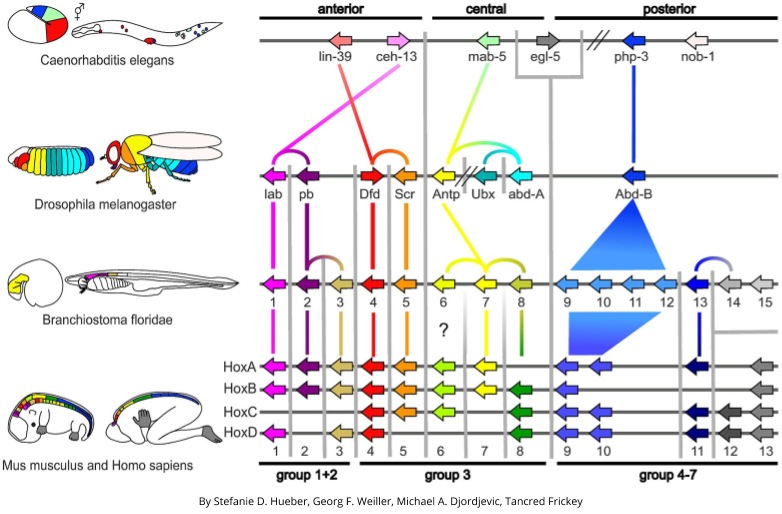
Hox genes help explain the phenomenon of Williston’s Law (module customization). In arthropod segments, boundaries in Hox gene formation promote customizations across different segments:
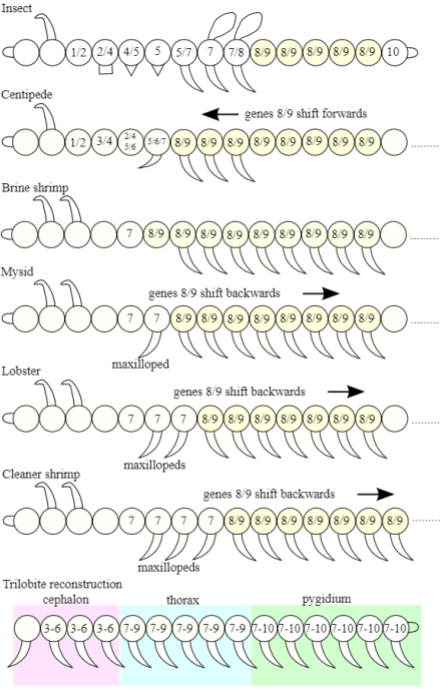
Bodybuilding & the Genetic Toolkit
In both flies and humans, the very same gene (Pax-6) orchestrates eye development, despite enormous differences in eye phenotypes. Even if you activate this gene in the wing of a fly, that wing will grow eye tissue. When the gene is deactivated, eye formation fails. And if you transplant the fly’s Pax-6 gene into a eyeless mouse, that mouse will regain the ability to grow its eyes.
The Pax-6 gene is an example of a master bodybuilder gene. Here are two other examples from this category
- The DLL “Distal-Less” gene builds appendages. In chickens it builds legs; fins in fish, siphons in sea squirts, and tube feet in sea urchins.
- The NK2 “Tinman” gene contributes to the circulatory system. It orchestrates heart development across many different phyla.
There is more to the story than just Hox and bodybuilder genes. Other “master” genes are shared across all animal phyla. These include genes for hormones, those that regulate cell type, those involved in signaling pathways, coloration, receptor mechanisms, and other DNA binding use cases. Together, these genes comprise the genetic toolkit: a set of genes responsible for the development of multicellular organisms.
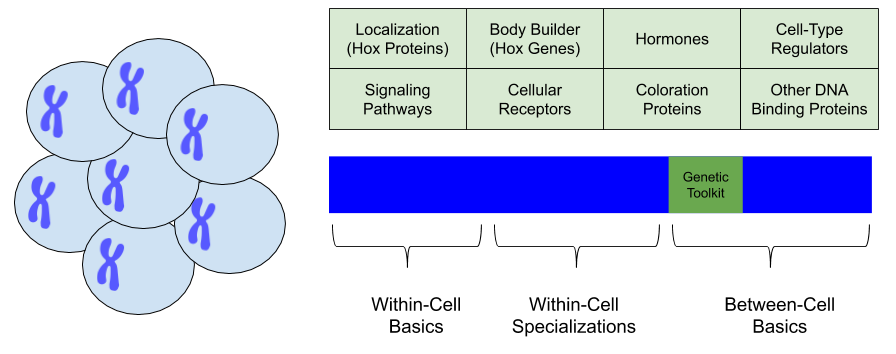
Explaining the Cambrian Explosion
Often two different species will have a feature in common. Such facts can be explained in two different ways.
- Homology: the feature is shared because it was invented in a common ancestor of both species.
- Homoplasy (aka analogy, or convergent evolution): the feature was not derived from a common ancestor; it was invented separately and independently
One example of homology is having four limbs: our tetrapod ancestors were the first to try this new body plan. In contrast, the evolution of wings in birds and bats is an example of homoplasy – the common ancestor of these species was terrestrial. This example is nicely illustrated in a phylogeny:

Protostomes and deuterostomes use the very same Hox and bodybuilder genes. It is very unlikely that the exact same genetic toolkit was constructed twice. The most parsimonious explanation is homology. Their common ancestor, a bilateral symmetric population Urbilateria, also possessed this genetic toolkit. Specifically, we can safely conclude that Urbilateria had a toolkit if at least six or seven Hox genes, Pax-6, Distal-Less, Tinman, and a few hundred more bodybuilding genes.
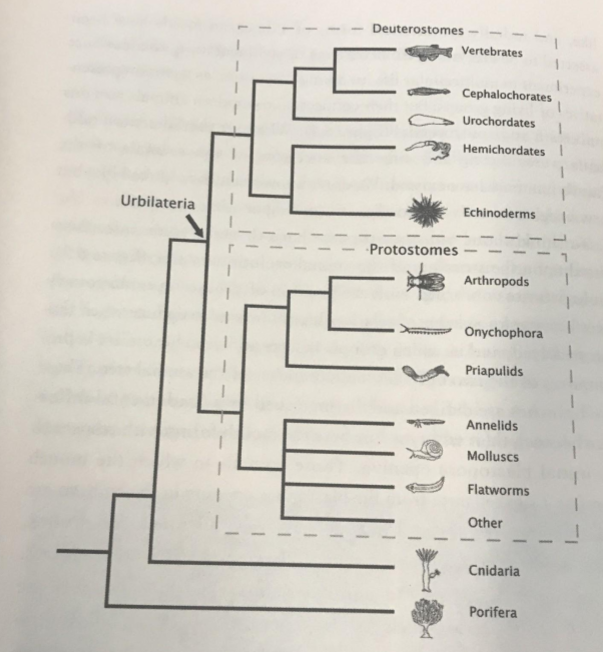
Urbilaterians have not yet been found in the fossil record. However, we can do better than envisioning some featureless worm. We can use our knowledge of their genome to infer their body plans.
Because Pax-6 resides in both branches of bilaterians, Urbilateria probably had some kind of light-sensing organ. Similar inferences from homologous genes adds more detail to this portrait. The first bilateral population probably had some form of appendage, a primitive heart, a through-gut with mouth and anus, and a diverse set of cell types (including photoreceptive, nerve, muscle, digestive, secretory, phagocytic, and contractile).
One of the great mysteries of evolutionary biology is the Cambrian Explosion, an adaptive radiation where dozens of new phyla appear in the fossil record in the span of about 40 million years. The genetic toolkit was fully in place by the time of the Cambrian Explosion. It seems likely that the compilation of the toolkit was a important prerequisite for such a radiation (although ecological factors surely also played a role).
Deep Homology
Consider the evolution of the eye. Evolutionary biologists once thought eyes had evolved independently dozens or even hundreds of times. This remarkable feat of evolution was attributed to strong selective pressure: having a light-sensitive organ just pays off, and the selective pressure is overwhelming enough to induce many species towards the same end product.
But modern genomics has revealed that these “independent inventions” actually derive from the redeployment of the conserved Pax-6 gene. The diversity of modern eyes is the result of specializations built on top of this basic genetic framework.
More generally, the deep homology hypothesis suggests that the body organization of all bilaterians derives from a substantial swathe of genes that comprise our genetic toolkit. Bilaterians do not invent novel developmental regimens whole-cloth. Rather, once the full toolkit assembled, changes in phyla occurred via alterations of regulatory circuits.
This principle sharpens how scientists explore new hypotheses. For example, humans are unique among primates for our penchant for vocal mimicry (we can learn how to produce novel sounds). But our more distant relatives (parrots, even seals) practice vocal mimicry. Because so much of our genetic material is conserved, we cannot afford to ignore similarities with even our distant relatives.
Until next time.












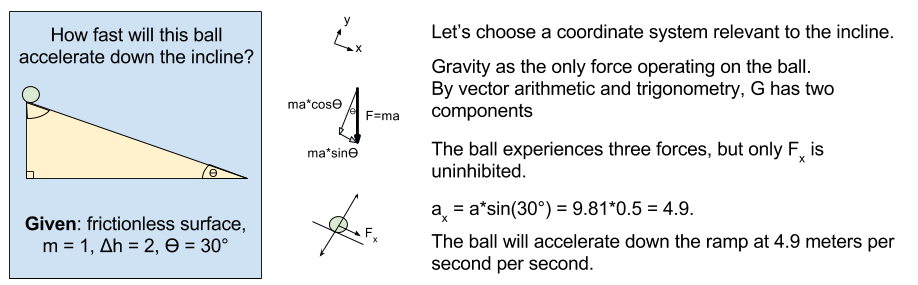
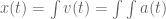
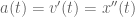

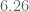 meters per second
meters per second 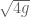 m/s). However, recall the classical definitions of kinetic and potential (gravitational) energy, which are
m/s). However, recall the classical definitions of kinetic and potential (gravitational) energy, which are 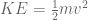 and
and  .
.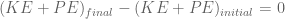
![m[(0 + 2g) - (\frac{ \sqrt{(4g)^2} }{2})] + 0 = m[2g - 2g] = 0](https://s0.wp.com/latex.php?latex=m%5B%280+%2B+2g%29+-+%28%5Cfrac%7B+%5Csqrt%7B%284g%29%5E2%7D+%7D%7B2%7D%29%5D+%2B+0+%3D+m%5B2g+-+2g%5D+%3D+0&bg=ffffff&fg=555555&s=0&c=20201002)
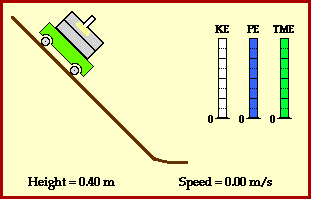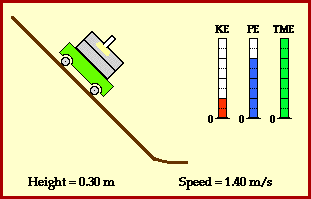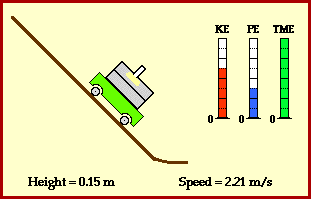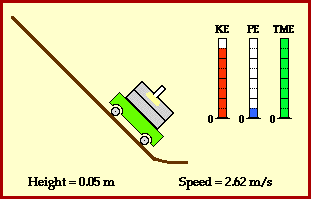
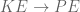 ? Because it is experiencing a force.
? Because it is experiencing a force.