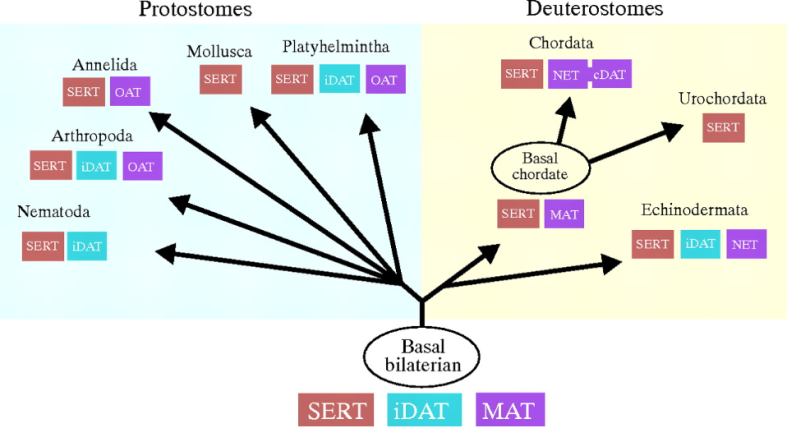
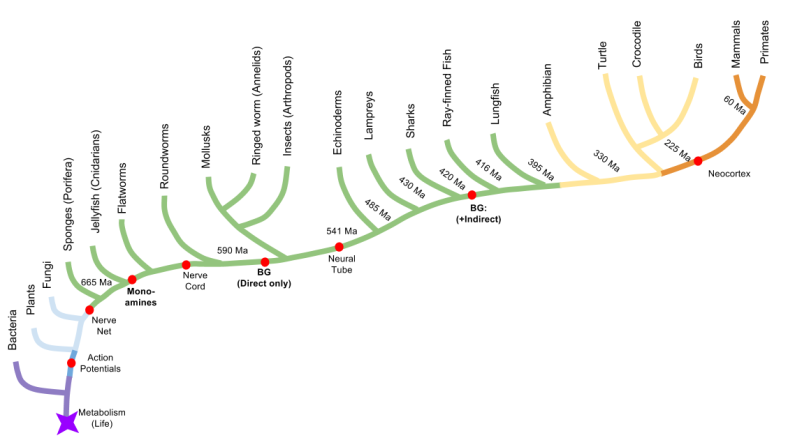
Part Of: Affective Neuroscience sequence
Content Summary: 800 words, 8 min read
Primary Emotion
There are many possible emotions. How can we make sense of this diversity?
Primary emotions are often used to shed light on our emotional lives. Like primary colors, these emotions blend together to reconstitute the full spectrum of emotional experience. For example, contempt is viewed as a combination of anger and disgust.
An emotion qualifies as primary if it satisfies the following criteria:
- Unique Machinery. It must be localized to specific neural processes.
- Known Signature. A fixed set of phenomenological and behavioral expressions
- Universal (Pre-Cultural). Expressed in all members of a given species. For ecologically valid stimuli, response does not detract from overall fitness.
- Primitive (Pre-Cognitive). Activated more strenuously during early development or immediate crisis (i.e., with minimal cognitive regulation).
- Differentiable. Can be dissociated from other primary emotions.
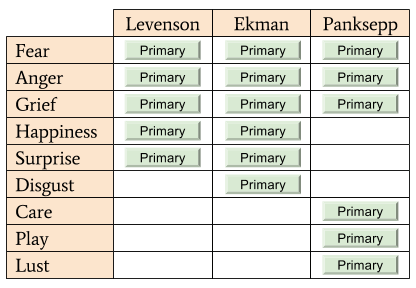
Despite consensus about the above criteria, there is less agreement on which emotions deserve membership. Here are three representative lists.
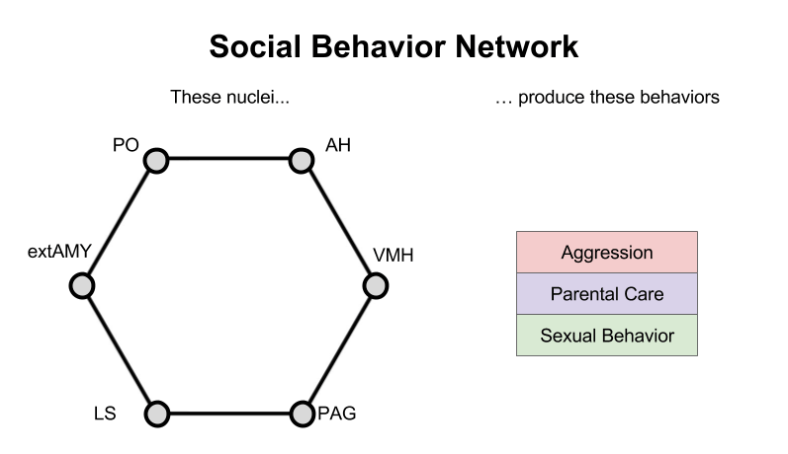
The Social Behavior Network (SBN)
Neuroscientists studying aggression have identified six brain regions that seem to produce this behavior. They are:
- Preoptic Area of the Hypothalamus (PO)
- Anterior Nucleus of the Hypothalamus (AH)
- Ventromedial Nucleus of the Hypothalamus (VMH)
- Periacquductal Gray (PAG)
- Lateral Septum (LS)
- Extended Amygdala (extAMY)
If any of these regions are damaged, an animal often becomes less aggressive. If you electrically stimulate these regions, the animal becomes enraged.
What is interesting about these six regions is that they were independently discovered by other neuroscientists who labelled them as the seat of parental care.
… AND, by yet other neuroscientists who had been investigating the neural basis of sexual behavior.
What do { Parental Care, Aggression, Sexual Behavior } have in common? They are entirely directed at members of one’s own species. These primary emotions are deeply related to animal social behavior.
Since the six nuclei { PO, AH, VMH, PAG, LS, extAMY } contribute to each of these three emotions & behaviors, they are now called the social behavior network (SBN).
Will it turn out that all social primary emotions are created by the SBN? I don’t know. It is suggestive, however, that Play has been partially localized to the lateral septum (LS).
SBN and Emotion Selection
The SBN is one brain structure that can produce three distinct emotional response. How is this possible? How does each emotion individuate itself within a single apparatus?
To proceed, we consult our “theorizing roadmap”:

Conceptually, we are plagued by “too many emotions”. Thus, we can either:
- Examine whether our three emotions can be unified; or
- Look for granularity within the SBN
Since the former is impractical, let’s look more carefully at the SBN.
One way to explain emotion individuation would be a shape hypothesis. If the intensity of neuron firing is encoded by height, you might expect different topographies (landscapes) to encode different emotions.

Another hypothesis is the granularity hypothesis. This posits that there may be e.g., three subdivisions of the lateral septum, and each subdivision supports a different emotion.

I tend to find this approach more plausible, given my experience with other subcortical structures. That said, time will tell. 🙂
Relation To The Basal Ganglia
The SBN is anatomically related to the basal ganglia. Recall that the basal ganglia has three loops: Associative, Sensorimotor, and Limbic. The SBN is strongly connected to, and shares two nodes with, the Limbic Loop.
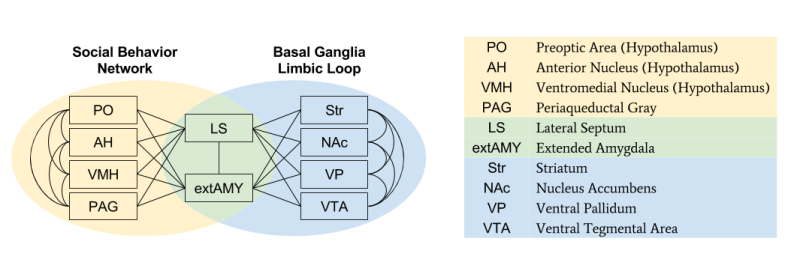
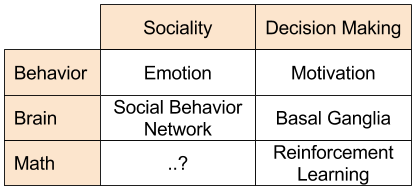
As we have seen, the basal ganglia is the seat of motivation. The anatomical connection between SBN and basal ganglia mirrors the behavioral link between sociality and motivation. However, on a mathematical level, it is less clear how social emotions can be incorporated into the reinforcement learning apparatus:
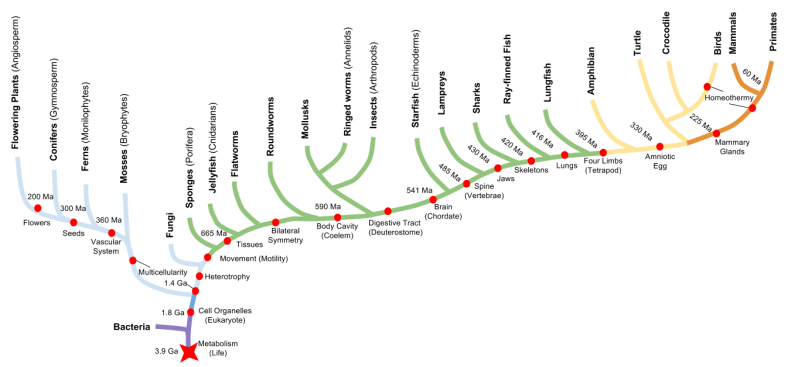
Evolution of Emotion
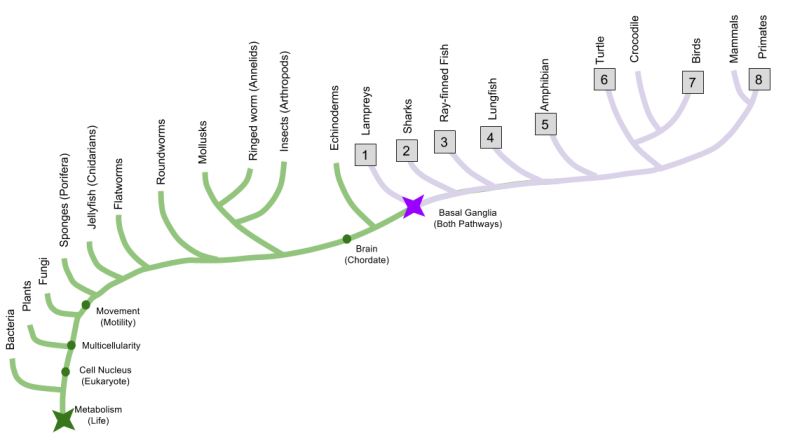
Let’s use comparative anatomy to discover when the social behavior network evolved. By dissecting brains from five representative species, we can infer that the basal ganglia dates back to at least the origin of ray-finned fish.
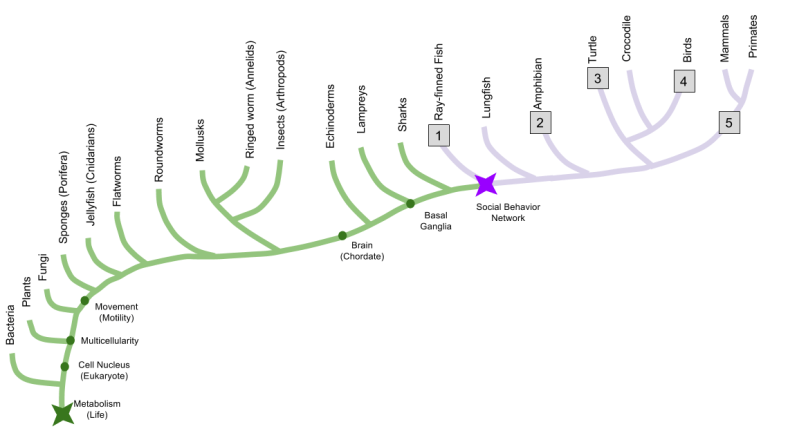
The SBN nuclei are preserved across our representative species:
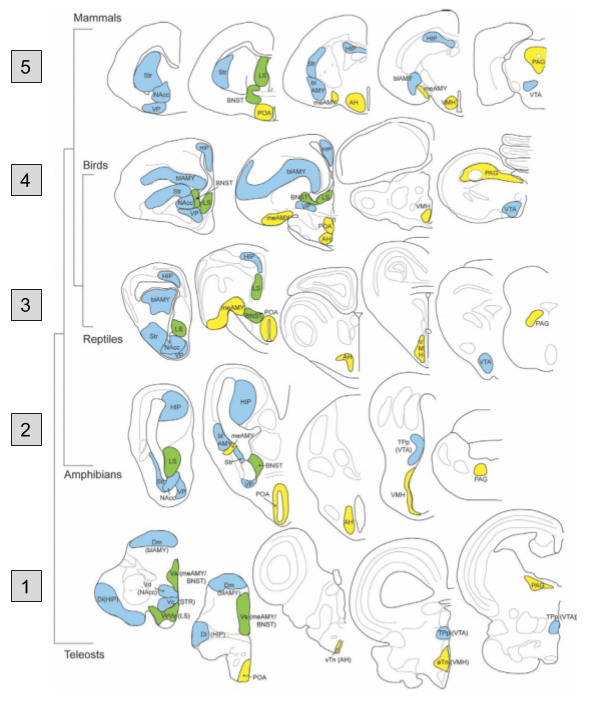
And hodology (connections) between SBN nuclei are preserved:
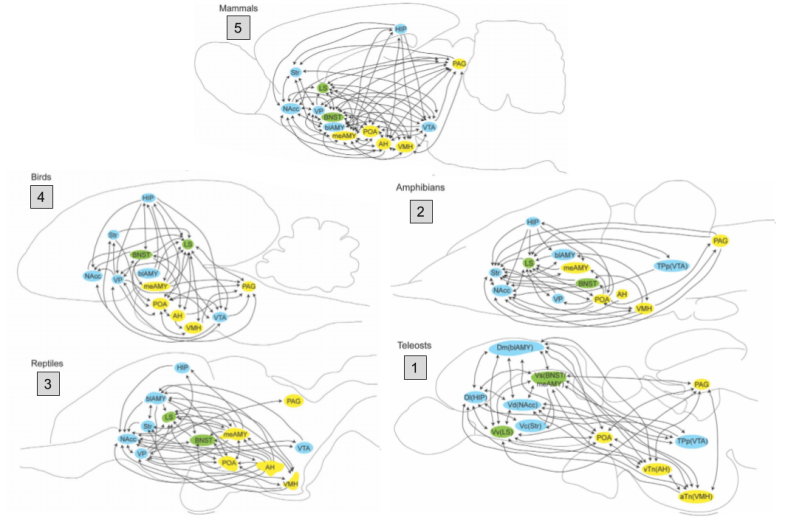
This evidence demonstrates that the social behavior network has been around since the invention of vertebrates. It also raises important questions, such as:
- How has the SBN changed to support hyper-social animals like primates?
- How much further back do emotional adaptations go? Do insects feel emotions? If yes, which kinds?
Until next time.
Related Works
- Newman (1999). The Medial Extended Amygdala in Male Reproductive Behavior: A Node in the Mammalian Social Behavior Network
- O’connell, Hofmann (2011). The vertebrate mesolimbic reward system and social behavior network: a comparative synthesis