
Part Of: Affective Neuroscience sequence
Followup To: An Introduction to Homeostasis
Content Summary: 2800 words, 14 min read
Living, Fast and Slow
We have previously learned how the brain engages in biological defense of physiological balance points conducive to life. For example, temperature is regulated by deploying both internal effectors (e.g., shivering, goose flesh, vasoconstriction) and external effectors (i.e., seeking warmth in the environment).
On this view, a stressor is anything in the outside world that knocks you out of homeostatic balance, and the stress-response is what your body does to reestablish homeostasis.
What happens when the organism is severely stressed? Hunting your prey, being hunted, fighting for females, freezing to death, and having sex each levy unique behavioral demands. But they also require many things in common:
- Sharpened attention & faster reflexes
- High heart rate & blood pressure to enhance blood supply to the organs.
- More energy (more glucose in the bloodstream, intracellular anabolism) to subsidize rapid movement.
- Pausing long-term projects (e.g., digestion consumes 10-20% of the mammalian energy budget)
What happens when an organism is comfortable? We might predict these effectors to be offline, and bodily resources to be engaged in projects with long-term benefits: sleep, digestion, cell repair, reproductive activity, protein construction (catabolism) etc.
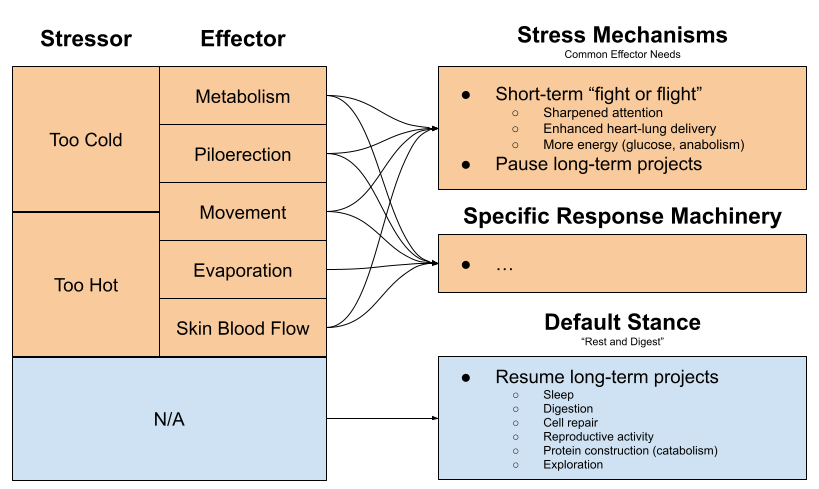
The stress response is not identical to physiological actions required to sustain life. But it is the embodiment of overlapping needs within the effector ecosystem. Call this the effector overlap theory of stress. The generality of the stress response explains why its discoverer Selye dubbed it the General Adaptation Syndrome.
The above account distinguishes between short-term emergencies versus long-term projects. On the life history theory of stress, stress involves postponing long-term projects. We can see this in semelparous species like salmon or which evolved to die immediately after reproduction. These salmon die from a prolonged stress response. Remove their overactive adrenal gland, and they are suddenly able to live for another year.

We previously distinguished between reactive vs predictive homeostasis. On the overlap theory, it should not be surprising that the stress response also exhibits a predictive, circadian component.
The Anatomy of Stress
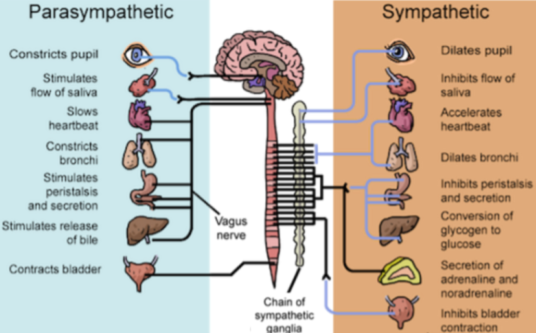
This basic dichotomy manifests in the parasympathetic (PNS) “rest and digest” and sympathetic (SNS) “fight or flight” systems. Nearly every organ is dually innervated by both systems, with the vagus nerve carrying most parasympathetic signals, and the sympathetic trunk mediating sympathetic signals. Moreover, these two subsystems often constitute opponent processes; that is, they exhibit functional antagonism.
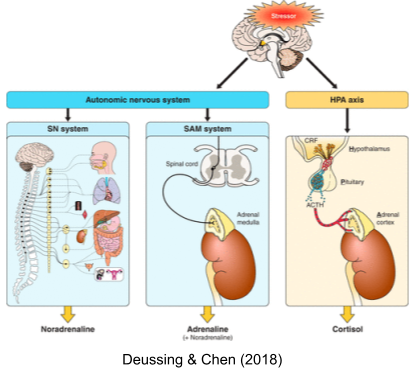
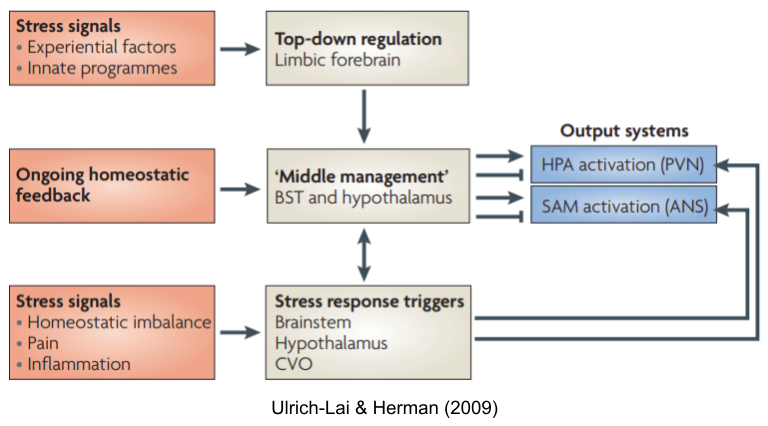
But the stress response involves more than just the SNS. We have previously introduced circumventricular organs (CVOs), which puncture the blood-brain barrier in a controlled way. These organs provide neuroendocrine integration. There are many neuroendocrine axes, including HPG (reproduction), HPT (metabolism) and HPS (growth). The hypothalamus-pituitary-adrenal (HPA) axis also regulates levels of glucocorticoids (cortisol in humans). Stressors also evoke other hormonal responses, including
- glucagon (energy mobilization),
- prolactin (suppressing reproduction),
- endorphins (blunting pain perception), and
- vasopressin (mediating the cardiovascular stress response)
- osteocalcin (inhibits the parasympathetic branch) Berger et al (2019).
Another branch coordinates activity between SNS and HPA: the sympathomedullary (SAM) pathway.
The affective dimension of arousal involves increased alertness to sensory stimuli, increased motor activity and increased emotional reactivity (Pfaff, 2006). Levels of arousal typically vary in a circadian fashion, but various events, including exposure to a stressor, can rapidly increase arousal levels. The orexin system appears to mediate the relationship between stress and arousal in the brain (Berridge et al 2010).
In periphery, norepinephrine (NE) and acetylcholine (ACh) mediate the SNS and PNS, respectively. In the brain, loosely speaking, these same neuromodulators are associated with arousal and learning. A good example of core-periphery functional consilience.
Taken together, these systems (SNS, HPA axis, SAM, etc) are known as the stress response system (SRS).
Stress Signatures
There are two kinds of stress response
- Interoceptive (systemic) stress, where the body detects homeostatic imbalance. Related to reactive homeostasis.
- Exteroceptive (psychogenic) stress, or predicted future dysregulation. Related to predictive homeostasis.
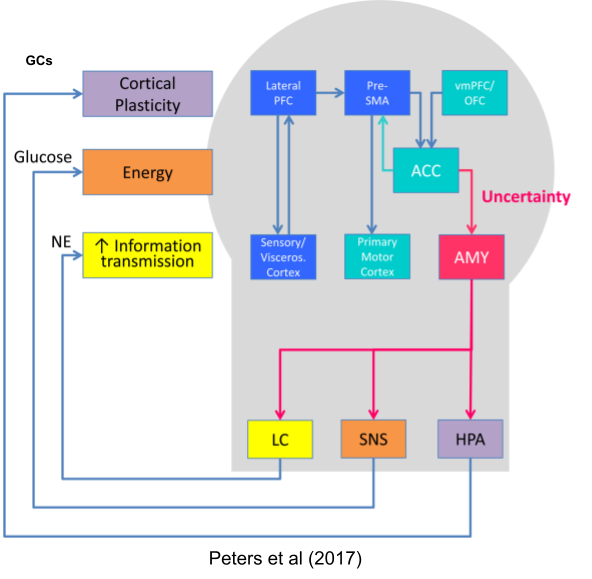
These forms of stress are handled differently by the brain,
There are two kinds of stress responses:
- Bodily stress response (e.g., physical exercise). Mediated primarily by noradrenaline via the SNS system.
- Mental stress response (e.g., working memory tasks). Mediated primarily by adrenaline via the SAM system.
Many interoceptive stressors require bodily responses, whereas some exteroceptive stressors only require mental responses.
The Experience of Stress
Our experience frequently toggles between these two systems. As Sapolsky (2004) writes:
If you are a growing kid and you have gone to sleep, your parasympathetic system is activated. It promotes growth, energy storage, and other optimistic processes. Have a huge meal, sit there bloated and happily drowsy, and the parasympathetic is going like gangbusters. Sprint for your life across the savanna, gasping and trying to control the panic, and you’ve turned the parasympathetic component down.
The interplay between the autonomic nervous system is visible in respiration. The PNS is dominant during exhalation, and stress response is more active during inhalation. This may explain why exhalation-focused breathing (with low inhalation/exhalation ratio) can promote calmness.
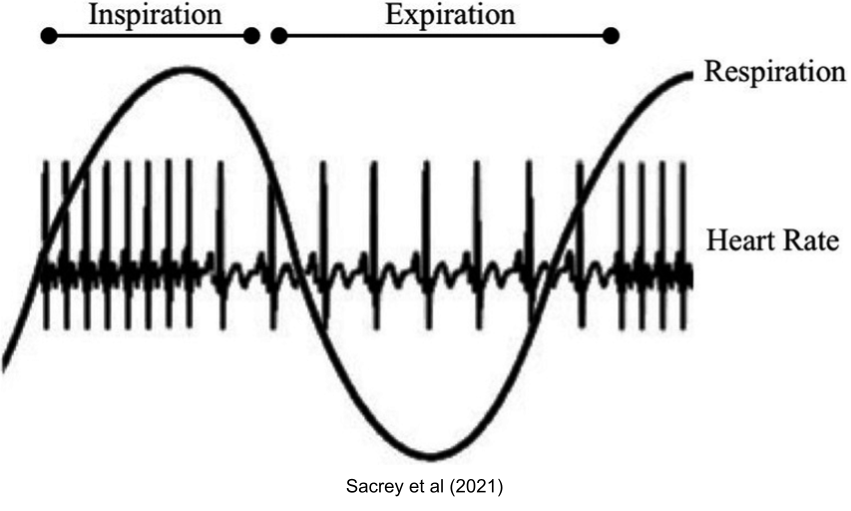
The interface between SNS and PNS is also well described during sexual intercourse:
To get an erection, a guy has to be calm, vegetative, and relaxed. What happens next, if you are male? You are having a terrific time with someone. Maybe you are breathing faster, your heart rate has increased. Gradually, parts of your body are taking on a sympathetic tone. After a while, most of your body is screaming sympathetic while, heroically, you are trying to hold on to the parasympathetic tone in that one lone outpost as long as possible. Finally, when you can’t take it anymore, the parasympathetic shuts off at the penis, the sympathetic comes roaring on, and you ejaculate.
The pain system is a competition between fast pain and slow pain (Hopkin 1997). Only the former promotes the stress response – they motivate you to quickly move away from the source of the piercing pain. What the slow fibers are about is getting you to hunker down so you can heal.
Stress can also cause analgesia (pain desensitization, e.g., a runner’s high), because your body secrets beta endorphins which mitigate objective pain perception in the spinal cord (Guillemin et al 1977). But when the stress response tilts towards anxiety, stress-induced hyperalgesia can occur via promoting subjective pain perception (Price 2000).
Heart attacks are much more likely to occur when the SRS is activated. So are flare-ups from autoimmune diseases like multiple sclerosis. Withdrawal and excessive drug use can be blocked by CRF antagonists, which speak to the intimate relationship between addiction and stress.
Three Phases of the Stress Response
These systems work at different timescales. SNS responds to stressors immediately, but cortisol changes occur within a few minutes. One can usefully model three phases of the stress response: stress initiation (high CRH, low cortisol), stress maintenance (high CRH and cortisol), and stress recovery (low CRH, high cortisol).
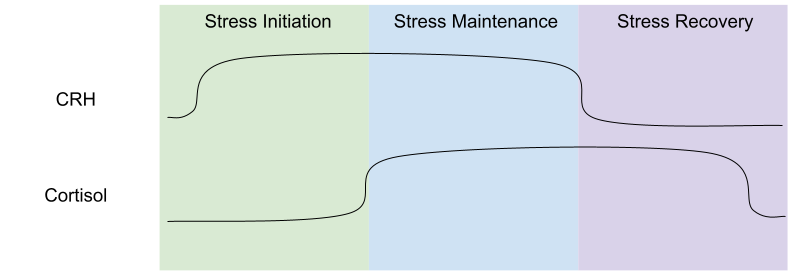
CRH inhibits eating (hypophagia), but cortisol promotes it (hyperphagia). The phase model of stress helps us make sense of this seemingly inefficient tension: eating is inhibited during the first two phases (high CRH), but promoted during stress recovery (low CRH). This explains why people exposed to long, continuous stressors (inhabiting the middle phase) tend to lose weight; whereas people with frequent, intermittent stressors (spending more time in recovery phase) tend to gain weight.
For those reactive to stress, stress eating can cause problems. Cortisol hypersecretors are most likely to be hyperphagic after stress (Epel et al 2001), and they disproportionately favor comfort foods (Dallman et al 2003). These people are more likely to have visceral fat (Epel et al 2000), which is much more harmful than subcutaneous fat (Welin et al 1987). This is because glucocorticoids are disproportionately expressed in the abdomen (Rebuffe-Scrive et al 1990), and are activated during stress recovery.
A similar trade off occurs in the immune system. The beginnings of stress response (first 30min) promotes immune function, but after a while (past 1h mark) stress has an immunosuppressant effect. This explains why long, continuous stressors suppress the immune system; whereas frequent, intermittent stressors stimulate the immune system and over time increases the risk of autoimmune disorders (e.g., asthma or multiple sclerosis). This explains why putting people “on steroids” (giving them massive amounts of cortisol) can protect against autoimmune disorders, and yet flare-ups of autoimmune symptoms are also yoked to stress.
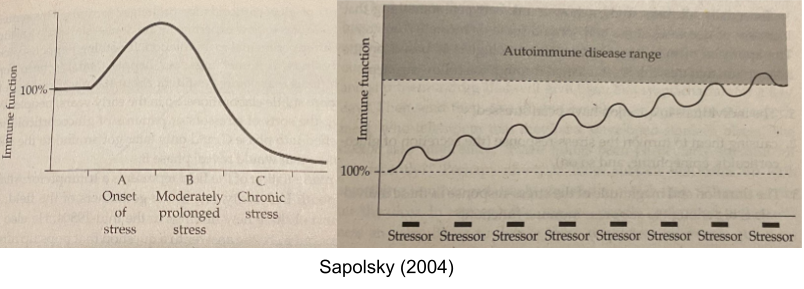
Herpesviruses establish latency, which means they hide inside cells and only replicate when they are at an advantage. Their DNA contains a glucocorticoid sensor – when the virus detects elevated GCs, it knows the immune system is temporarily suppressed & it comes out of latency. The virus can even artificially induce an SRS response via your hypothalamus.
Chronic Stress and Senescence
While today’s discussion is at the systems level, the SRS also interacts with intracellular mechanisms. As we will explore next time, individual differences in chronic stress may involve differential rates of senescence. For example, individuals exposed to chronic stress show signs of accelerated biological aging, such as telomere erosion (Humphreys et al 2012).
Many mechanistic theories of senescence appeal to metabolic functions of the mitochondria. And chronic stress has been shown to reduce mitochondrial energy production capacity (Picard & McEwen 2018).
The reactive oxidative species (ROS) theory of senescence appeals to intracellular oxidative stress, not to be confused with systemic psychological stress. But chronic psychological stress does seem to promote oxidative stress (Aschbacher et al 2013), which may explain its role in accelerated aging.
Stress reactivity (larger, slower-fading GC response to stressors) is caused by chronic stress. The Big-5 personality trait of Neuroticism is a crucial moderator of reactivity (Zobel et al 2004)
The Volatility facet of Neuroticism, analogous to Hostile Type A personalities, produces an increased risk of cardiovascular disease (CVD) (Williams & Litman 1996). The link seems causal (Friedman et al 1996). Amusingly, the link between Type A and CVD seems to have been first discovered by an upholsterer in a cardiologist’s office: “…what on earth is wrong with your patients?”:

Stress is involved in age-related diseases like general inflammation, and CVD. However, GC seems to not play a causal role in another age-related disease: cancer. Another endocrine system implicated in senescence, insulin-like growth factor-1 (IGF-1), is more involved in cancer risk.
Stressor Determinants: Predictability and Control
Gradual-onset ulcers are caused by an acid-resistant bacteria known as helicobacter pylori (Dooley & Cohen 1988). Yet, while nearly all of us have the disease, only 10% of us develop ulcers. Stress is one of the lifestyle factors which inhibits repair of the stomach lining, and hence increases risk of ulceration (Levenstein 1998). Ulceration is thus a useful operationalization of stress, and you can explore intricacies of the SRS by measuring gastric lesions.
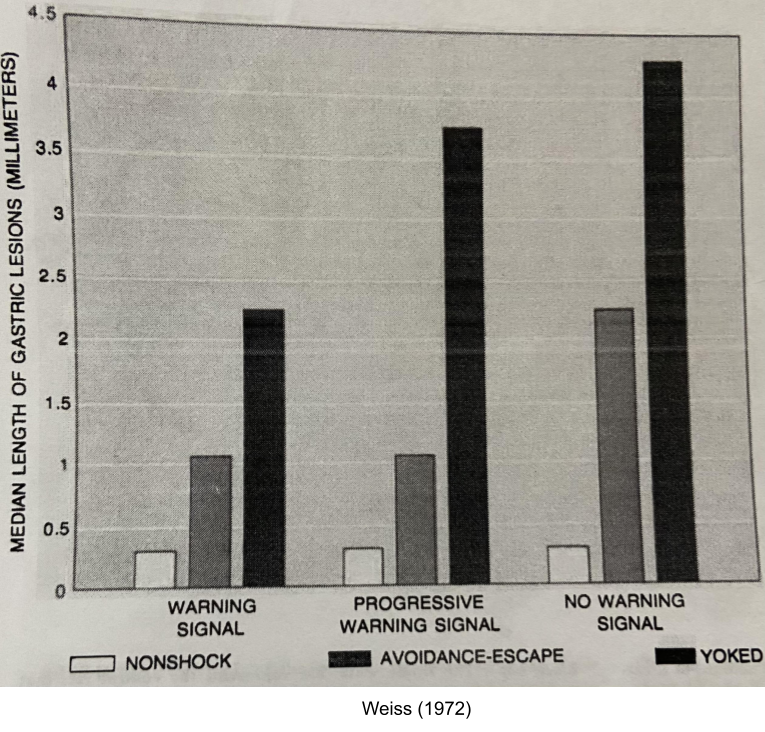
Weiss (1972) hooked mice to receive tail shock every minute in a continuous 21-hour session. His work sheds light on psychological mechanisms of coping.
- Predictability: if you give a warning tone before the shock, rats experience less stress (left vs right column)
- Control: give a lever to stop its shock and a “yoked” neighbor, the rat will experience less stress (gray vs black bar).
- Foreknowledge: early foreknowledge of shock worsens stress only in rats with no control (left vs middle column).
Predictability can drive SRS magnitude, independent of the physiological implications:
- Random interval feeding is associated with stronger SRS response than fixed intervals in rats.
- Stressor habituation reduces the response to predictable stressors. Parachute jumpers eventually experience no stress as they move through their training (Ursin et al 1978).
- In WW2, nightly air raids in cities produced fewer ulcers than infrequent & unpredictable suburban bombings (Stewart & Winser 1942).
Uncontrollability can shut down the SRS. Overmeier & Leaf (1965) discovered that dogs exposed to uncontrollable stressors were later unable to learn a shock-avoidance task, one that controls were easily able to master. Such learned helplessness has tight links to major depression (Seligman 1975). Polyvagal theory interprets this as Jacksonian dissolution: if the SRS fails, the nervous system will fall back to the dorsal vagus.
White (1959) described the desire for control as a motivational drive for competence. As behaviorism was supplanted during the Cognitive Revolution, Rotter (1966) reformulated the theory as locus of control: attitudes of self-control are associated with positive outcomes in myriad facets of daily life (Lefcourt 1992). People’s ratings of self-efficacy are better predictors of future behavior than their past behaviors (Bandura 1977). Self-efficacy also controls effort. When faced with difficulties, people who doubt their abilities quickly give up; people who don’t ratchet up their effort (Bandura & Cervone 1983)
These led to the attributional reformulation of learned helplessness; with its three parameters of explanatory style:
- Locus: is the cause internal to the self, or external?
- Consistency: are the causes stable over time, or not?
- Scope: are the causes global, or specific?
A person with a depressive style (habitually invoking internal, stable, and global factors to explain failures) is most at risk of becoming depressed in the face of uncontrollable circumstances.
Outlets for Frustration
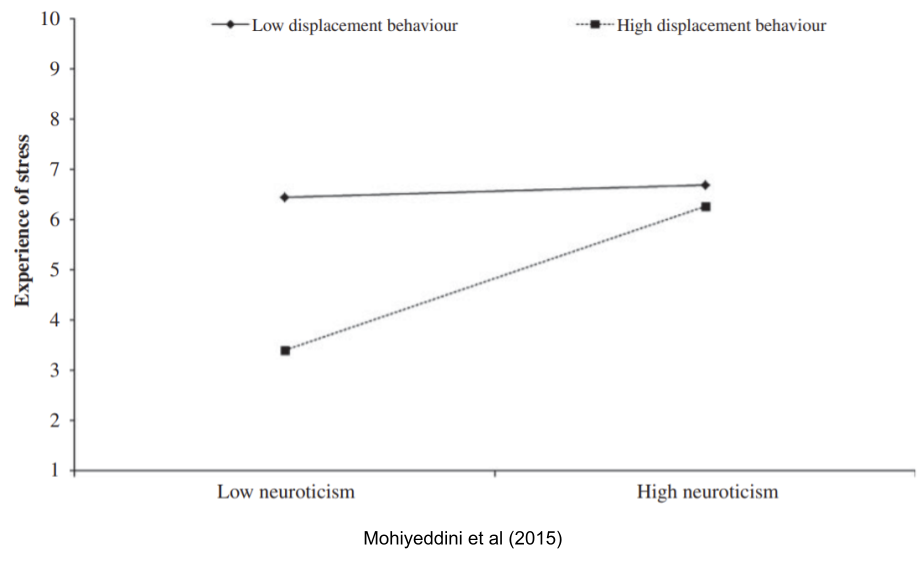
We have previously discussed displacement behaviors: for example, if a starving rat is given some food, but not enough to mitigate the drive, it will engage in ritualistic behaviors, such as pacing, gnawing wood, or self-directed behaviors. These behaviors reduce the stress response, but only for men with low neuroticism (Mohiyeddini et al 2015). Displacement behaviors don’t seem to help women in the same way (Mohiyeddini et al 2013).
Displacement aggression (“punching down”) is a significant outlet for frustration. Here’s Sapolsky (2004):
A variant of Weiss’s experiment uncovers a special feature of the outlet-for-frustration reaction. This time, when the rat gets the identical series of electric shocks and is upset, it can run across the cage, sit next to another rat and.. bite the hell out of it. Stress-induced displacement of aggression: the practice works wonders at minimizing the stressfulness of a stressor. It’s a real primate specialty as well. A male baboon loses a fight. Frustrated, he spins around and attacks a subordinate male who was minding his own business. An extremely high percentage of primate aggression represents frustration displaced onto innocent bystanders. Humans are pretty good at it, too. Taking it out on someone else–how well it works at minimizing the impact of a stressor.
I wish that I understood the reason why outlets have such effects.
Until next time.
References
- Abramson et al (1978). Learned helplessness in humans: critique and reformulation
- Aschbacher et al (2013). Good stress, bad stress and oxidative stress: insights from anticipatory cortisol reactivity
- Bandura (1977) Self-efficacy: toward a unifying theory of behavioral change
- Bandura & Cervone (1983). Self-evaluative and self-efficacy mechanisms governing the motivational effects of goal systems.
- Berger et al (2019). Mediation of the Acute Stress Response by the Skeleton
- Berkman & Syme (1979). Social integration, social networks, social support and health
- Berridge et al (2010). Hypocretin/Orexin in Arousal and Stress
- Dallman et al (2003). Chronic stress and obesity: a new view of comfort food
- Deussing & Chen (2018). The Corticotropin-Releasing Factor Family: Physiology of the Stress Response
- Dooley & Cohen (1988). The clinical significance of Campylobacter pylori
- Epel et al (2001). Stress may add bite to appetite in women: a laboratory study of stress-induced cortisol and eating behavior
- Epel et al (2000). Stress and body shape: stress-induced cortisol secretion is consistently greater among women with central fat
- Friedman et al (1996) Effects of Type A behavioral counseling on frequency of episodes of silent myocardial ischemia in coronary patients
- Guillemin et al (1977). Beta-endorphin and adrenocorticotropin are secreted concomitantly by pituitary gland.
- Heinrichs et al (2003). Social support and oxytocin interact to suppress cortisol and subjective responses to psychosocial stress
- Hopkin (1997). Show me where it hurts: tracing the pathways of pain
- Humphreys et al (2012). Telomere shortening in formerly abused and never abused women.
- Lefcourt (1992) Durability and impact of the locus of control construct
- Levenstein (1998). Stress and peptic ulcer
- Marmot et al (1978). Employment grade and coronary heart disease in British civil servants
- Mohiyeddini et al (2013). Displacement behavior is associated with reduced stress levels among men but not women.
- Mohiyeddini et al (2015). Neuroticism and stress: the role of displacement behavior
- Overmeier & Leaf (1965). Effects of discriminative Pavlovian fear conditioning upon previously or subsequently acquired avoidance responding.
- Parent et al (2017). Dynamic stress-related epigenetic regulation of the glucocorticoid receptor gene promoter during early development: The role of child maltreatment.
- Peters et al (2017). Uncertainty and stress: why it causes diseases and how it is mastered by the brain
- Pfaff (2006). Brain arousal and information theory: neural and genetic mechanisms
- Picard & McEwen (2018). Psychological Stress and Mitochondria: A Systematic Review
- Price (2000). Psychological and Neural Mechanisms of the Affective Dimension of Pain
- Rebuffe-Scrive (1998). Steroid hormones and distribution of adipose tissue
- Rebuffe-Scrive et al (1990). Steroid hormone receptors in human adipose tissues.
- Rosengren et al (1993). Stressful life events, social support, and mortality in men born in 1933.
- Rotter (1966). Generalized expectancies for internal versus external control of reinforcement.
- Sacrey et al (2021). Slow-paced breathing: influence of inhalation/exhalation ratio and of respiratory pauses on cardiac vagal activity.
- Sapolsky (2004). Why zebras don’t get ulcers, third edition
- Seligman (1975). Helplessness: on depression, development, and death.
- Stewart & Winser (1942). Incidence of perforated peptic ulcer: effect of heavy air-raids.
- Ulrich-Lai & Herman (2009). Neural regulation of endocrine and autonomic stress responses
- Ursin et al (1978). Psychobiology of stress: a study of coping men
- Welin et al (1987). Family history and other risk factors for stroke: the study of men born 1913
- Weiss (1972). Psychological factors in stress and disease
- White (1959). Motivation reconsidered: the concept of competence
- Williams & Litman (1996). Psychosocial factors: role in cardiac risk and treatment strategies
- Zobel et al (2004). High neuroticism and depressive temperament are associated with dysfunctional regulation of the hypothalamic–pituitary–adrenocortical system in healthy volunteers
