
Part Of: Affective Neuroscience sequence
Content Summary: 2700 words, 13 min read
Three Behavioral Systems
Physiological variables are not just regulated by the viscera, but also by behavior. Such behaviors come in two categories:
- Approach behaviors, towards objects of positive valence (e.g., towards food)
- Avoidance behaviors, away from objects of negative valence. (e.g., away from predators)

You can see a similar dichotomy in AI systems:

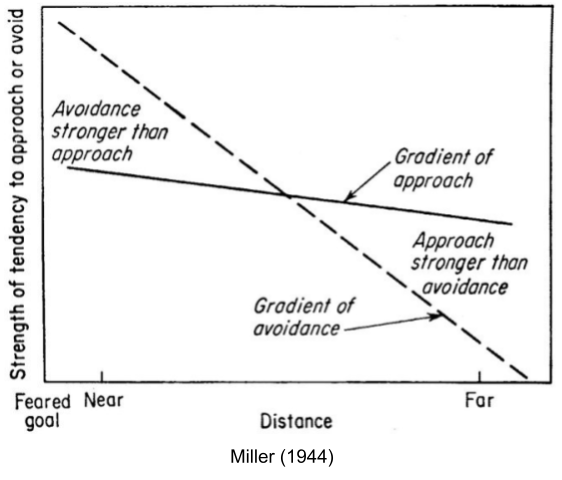
One might venture that approach and avoidance are produced by a unitary system. But early behaviorist experiments showed that the strength of avoidance decreased at a greater rate than that of approach (Miller 1944). If you present food and shock together in an alley at the appropriate volumes, the approach system will dominate… until the rat gets close enough for its avoidance system to counterbalance. The result: fidgeting. Drugs like sodium amylobarbitone can modify the location of the conflict point. These data suggest, distinct systems control approach and avoidance behaviors.
The stress response system (SRS) is not synonymous with “fight or flight”. As we noted previously, the SRS engages behavioral effectors to address challenges to homeostasis. Behavioral effectors include avoidance – but they also include approach (e.g., foraging). Let us call the distinction between approach vs avoidance the bivalent motivation theory.
What about anxiety? Several theorists have sought to locate anxiety within the avoidance system. But anxiety dissociates from fear. Panicolytic drugs, which mimic amygdala lesions, mitigate fear but not anxiety. Anxiolytic drugs, which mimic hippocampal lesions, mitigate anxiety but not fear (Blanchard et al. 1997).
Gray & McNaughton (2004) argue that in some cases (novel objects, ambiguous social contexts) can concurrently activate the approach and avoidance systems. Rather than risking a disorganized response, this conflict resolution system is activated, and performs two functions:
- To suppress prepotent behavior candidates generated by the approach and avoidance systems.
- To elicit risk assessment behaviors to disambiguate the situation. And indeed, anxiolytic drugs suppress risk assessment behaviors (Blanchard et al 2011).
We will be referring to these three systems often.
- The avoidance system is also called the fight, flight, or freeze system (FFFS).
- The approach system is also called the behavioral activation system (BAS).
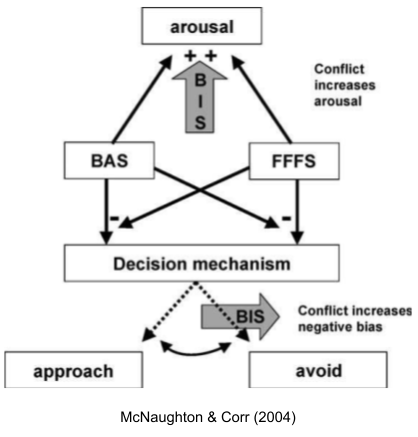
- The conflict resolution system is also called the behavioral inhibition system (BIS).
While much exploration occurs in a context of safety, BIS-mediated explorations are conducted in situations of high risk. Thus, the BIS promotes physiological arousal and the stress response, to prepare the body for a potential emergency. Finally, the BIS increases negative bias by sensitizing the FFFS avoidance system, which may reduce the risk of further wasteful approach-avoidance conflict.
Reactivity vs Coping Style
Koolhaas et al (2010) describe two coping styles: proactive coping (boldness) and reactive coping (shyness). These traits are consistently expressed in the same individual at different times and in different situations. For example, proactive individuals who response aggressively to resident intruders are more likely to produce defensive behavior in a burying test two weeks later.
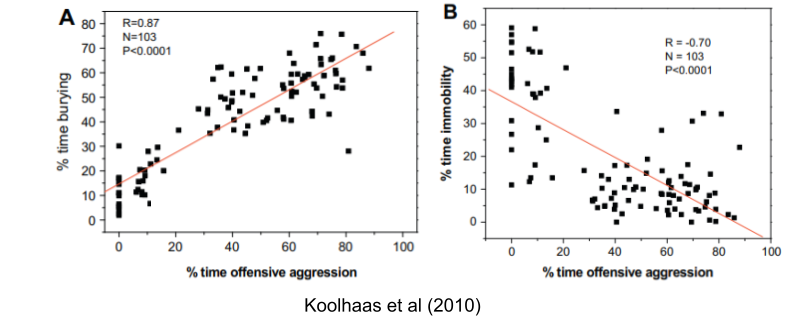
Reactive coping is also associated with individual differences in behavioral flexibility and impulse control! To me, this suggests reactive coping is analogous to an overactive BIS (although de Boer et al 2017 deny reactive coping relates to anxiety, nor the primary role of hippocampus).

Most well-known personality tests have poor statistical reliability… except one. The Big 5 (OCEAN) theory of personality explains large amounts of behavioral variation. Factor analyses reveal a hierarchy (DeYoung et al 2007):
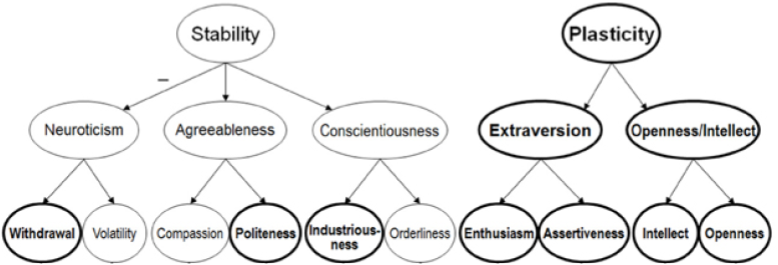
Neuroticism is associated with reactivity of the HPA axis. It has two major subfactors:
- Withdrawal reflects risk of anxiety and depression;
- Volatility a disposition towards irritability, anger, and emotional lability.
Koolhaas et al (2010) present a 2D model, with the HPA axis playing a role in reactivity, and serotonin playing a role in coping style. This nicely dovetails the personality constructs above.

Withdrawal (reactive coping) has been attributed to an overactive BIS system. In developmental psychology, this has been studied in children with behavioral inhibition (BI; see Kagan et al 1984), analogous to adult shyness (Barker et al 2018).
- High BI children produce a larger startle (Barker et al 2014), and enhanced amygdala activity (Cremers et al 2010): sensitized avoidance.
- High BI adults exhibit faster responses to both reward and punishment (Hardin et al 2006): sensitized approach (Helfinstein et al 2012).
Why does shyness occur? BIS only activates when the BAS and FFAS conflict. If both mechanisms expand, conflict occurs more frequently.
Evolution of Bivalence
These systems are extraordinarily ancient and conserved across phyla (Elliot & Covington 2001). In unicellular organisms, movement is dictated by gradients, such as foraging initiated by immediate chemical gradients in the immediate surroundings. While the representation capacities of these systems have clearly increased, the basic dichotomy persists.
Cisek (2021) proposes that bivalent motivation was a central organizing principle of neural evolution.

He also argues that
- Contralateral innervation evolved to support the avoidance system: if a predator is sensed by the left hemisphere, the animal needs to move to the right and vice versa.
- The approach system implemented winner-take-all decision-making, whereas the avoidance system is implements averaging. If you spot two predators, your escape vector should integrate both of them. But if you find two foraging patches, it is optimal to only approach one.
Biological Substrate
For the BAS approach system, tonic dopamine in the basal ganglia plays a central role in motivating approach. This is clearly seen in neurological conditions (Krack et al 2010):
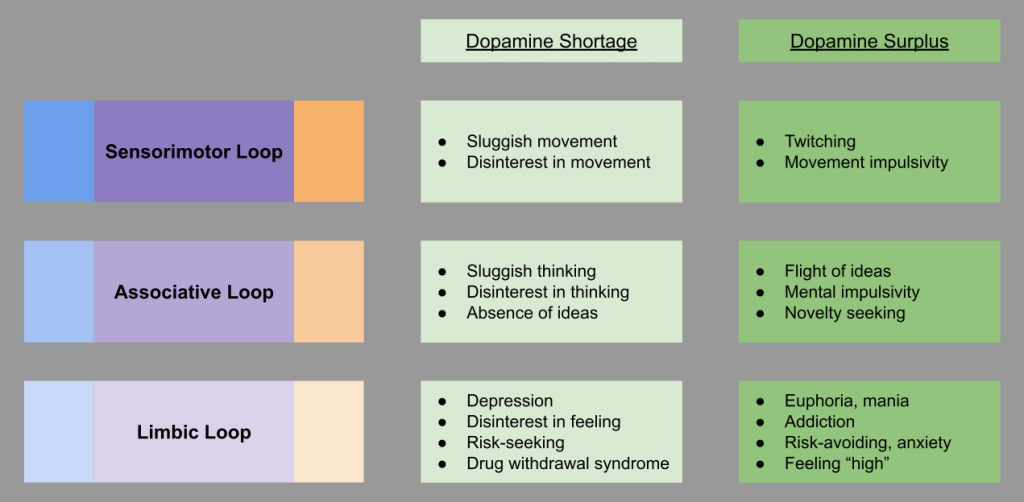
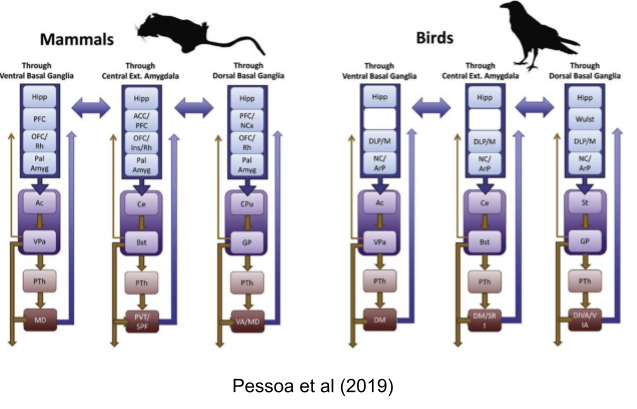
The FFAS avoidance system is grounded in the central amygdala and the bed nucleus of the stria terminalis (BNST). Just as the approach system calculates reward, the lateral habenula calculates antireward. This extended amygdala loop interoperates with the three “classical” basal ganglia loops above:
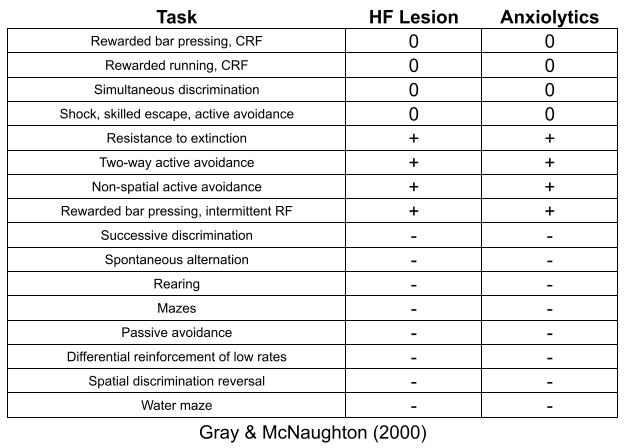
The BIS conflict detection system is closely associated with the septo-hippocampal system (Gray & McNaughton 1980). Anxiolytic drugs bear a strong resemblance to hippocampal lesions.
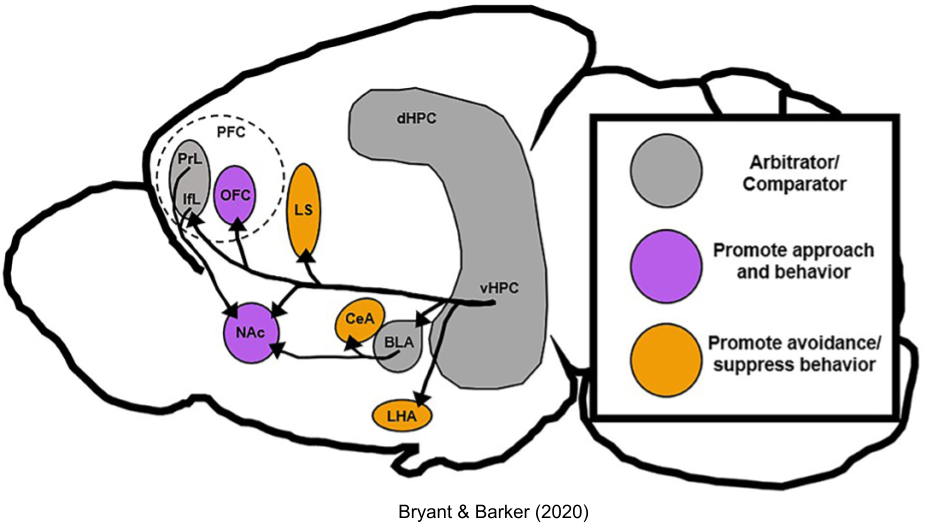
Of course, the cortex also plays an important role. Consider the pregenual cingulate (a.k.a. BA 25, or prelimbic cortex). This region has been implicated in depression (Pizzagalli 2011), addiction (Goldstein et al 2009), OCD (Fitzgerald et al 2005), and PTSD (Kasai et al 2008). While most of this surface had equal numbers of approach- and avoidance- representing neurons, one particular subregion (ventral pregenual anterior cingulate cortex, vPgACC) was predominated by avoidance processes.
Stimulation of vPgACC altered the decision boundary for approach-avoidance tasks, where the rat had to choose whether to experience food and an aversive airpuff, but not approach-approach tasks where the rat had to choose between two different kinds of food. These effects were cumulative: avoidance became increasingly pronounced as the number of stimulation trials increased. Finally, the effect of this region evaporated on administration of antianxiety drugs (Amemori & Graybiel 2012).

The striatum contains patches of cells (striosomes) in an otherwise homogenous population (matrix). The matrix appears to be only responsive to benefits; whereas the striosomes evaluates costs and benefits when both values are high. Striosomes in the dorsomedial associative striatum receive input preferentially from vPgACC, and their activity also changes the decision boundary by increasing loss aversion (Friedman et al 2015). Striosomes project gorgeous bouquets (Crittenden et al 2015) into the dopaminergic substantia nigra pars compacta (SNpc), which mediates their impact on decision making.
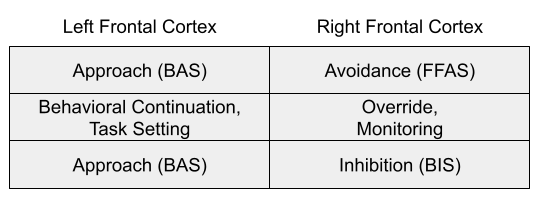
Lateralization in Frontal Cortex
In the cortex, some of the underlying mechanisms appear to be lateralized.
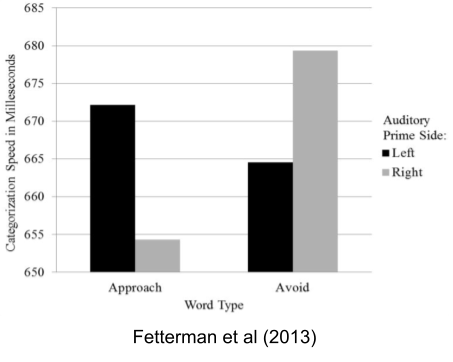
One tradition argues that the left frontal cortex participates in approach; the right in avoidance (Fetterman et al 2013; Carver & Harmon-Jones 2009).
- Chemical suppression (via amytal injection) to the left hemisphere produced depression; right injections produce mania. (Terzian 1964).
- Lesions to the left hemisphere produce depression, lesions to the right hemisphere produce mania (Robinson & Price 1982).
- Experiments which elicit reward processes create left-asymmetric activity; punishment created right-biased activity (Coan & Allen 2004)
- Personality differences in approach and avoidance are predicted by resting EEG asymmetries (Davidson 1999)
Another tradition argues that left vs right reflects behavioral continuation vs override (Wacker et al 2008), or task setting vs monitoring (Stuss & Alexander 2007). Both traditions seem close to convergence on the left hemisphere. I wonder if there might exist some middle ground, where the right hemisphere participates in the other avoidance system (BIS).
Right frontal theta is uniquely sensitive to approach-avoidance conflict, and is more pronounced in people suffering anxiety disorder (Shadli et al 2021). Such goal conflict-specific rhythmicity (GCSR) is now being used as a biomarker for anxiety disorder.
Consider posing asymmetries. The left side of the face is more active during emotional expression (Borod et al 1997). This explains why people prefer to show (and view) the left side of the face, and why people instructed to hide their emotions preferentially display the right.

The right hemisphere theory attributes perception & performance of all emotions to the right hemisphere. But there appears to be a second lateralization effect compatible with bivalent motivation (Demaree et al 2005). While on average the left hemiface is more expressive for all emotions, the left-bias was stronger for negative emotions, and weaker for positive emotions (Borod et al 1997).
Humans preferentially turn rightward in the (approach) act of kissing (Gunturkun 2003).
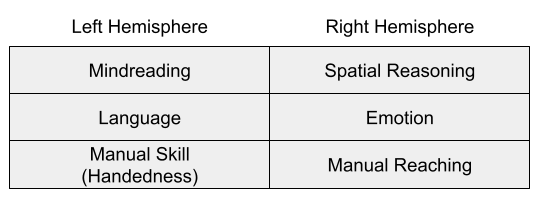
In general, left-hemisphere dominance for handedness and language might be compatible with a general specialization for routine action control (MacNeilage 1998), and can be reconciled with the postural origins theory of handedness (MacNeilage 2007).
The study of lateralization suffers several limitations:
- The shadow of mythology looms large (McManus 2018), exemplified in popular brainedness theories (e.g. Jaynes 1976, McGilchrist 2009).
- The comparative biology data of handedness is equivocal.
- The difference scores method in comparing hemispheric activity is contentious.
- These data often only permit macro-scale descriptions (“frontal cortex”), which inevitably conflate meso- and micro-scale structural circuits.
- I have not found a unifying theory to explain the relationship of lateralization effects across domains.
Yet these low-resolution data still suggest to me a left-bias for approach, and a right-bias for avoidance.
Hierarchical Organization
Stimulating avoidant nuclei in the hypothalamus produces fight, flight, or freezing. These behaviors depend not on the location of the probe, but on the context of the animal’s environment. Blanchard & Blanchard (1990) showed that considerable behavioral variation can be explained by defensive distance:
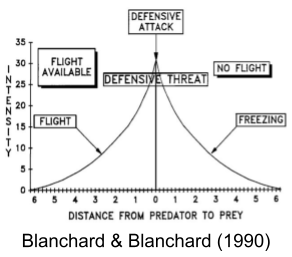
Defensive distance is more nuanced than physical distance. In a more dangerous situation, a greater real distance will be required to achieve the same defensive distance. Likewise, with a braver individual, a smaller real distance will be required to achieve the same defensive distance.
The behavioral hierarchy is driven by defensive distance. More distant threats produce slower, sophisticated defenses; more proximal threats produce faster, cruder defense mechanisms (Blanchard & Blanchard 1990). These are:
- Undirected escape (emotional correlate: dread)
- Directed escape (emotional correlate: panic)
- Active avoidance (emotional correlate: fear and phobias)
- Discriminated avoidance (which might include complex emotions such as guilt)
Recall the neural hierarchy: the further from the brainstem, the more conceptual the representations. Graeffe (1994) argues the behavioral hierarchy is isomorphic to a neural hierarchy, with phylogenetically newer structures layered on top of older systems. This is the hierarchical defense system.
The central amygdala supports fear conditioning (Le Doux 1994). Defense systems lower in the hierarchy do not support such learning, but directly respond to dangerous stimuli.
Just as BAS and FFFS are organized hierarchically, McNaughton & Corr (2004) argue the same for BIS:
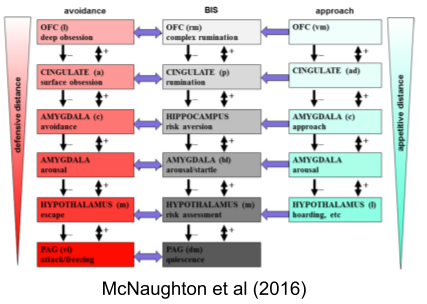
Economics and Emotion
It is important to distinguish between valuation and motivation. Specific visceral states dynamically alter individual valuations (this is alliesthesia). But even holding visceral deprivation constant, there are individual differences in valuation sensitivity. In behavioral economics, the ratio of positive to negative sensitivity is known as loss aversion.
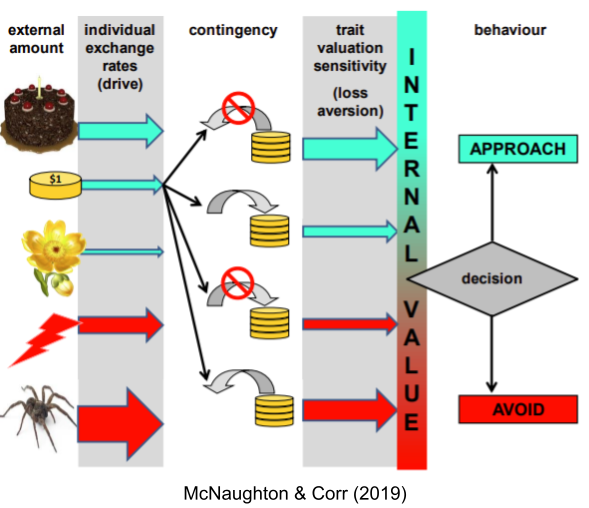
Loss aversion is not the same as repulsion sensitivity. Individual differences in valuation and motivation vary independently.
Modular theories of emotion (e.g Panksepp 1998) envision distinct neural circuitry for each basic emotion. In contrast, constructivist theories explore emotional states as emergent properties from a few core dimensions. The latter have been heavily influenced by Russel (1980) circumplex model, which organizes emotional experience into two dimensional space.
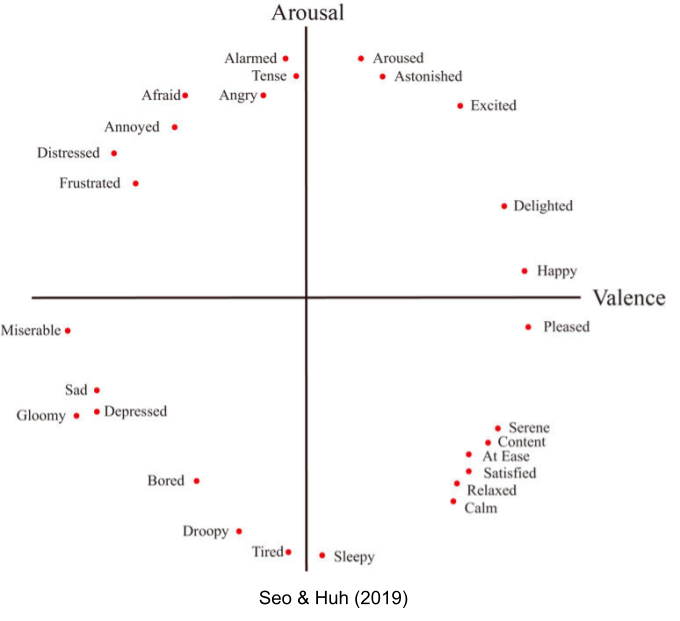
Approach/avoidance theorists are more aligned with dimensional models. But rather than interpreting valence as one-dimensional (bipolar), they claim each object is simultaneously attributed a positive and a negative valence score (bivalenced). Ambivalence is universal. Psychological evidence (van Harreveld et al 2004) and neural evidence (Man et al 2017) suggests brains retain multiple representations of valence. For example, the basolateral nucleus of the amygdala (BLA) contains three subpopulations of neurons: positive valence, negative valence, and arousal (Shabel & Janak 2009).
Until next time.
References
- Amemori & Graybiel (2012). Localized microstimulation of primate pregenual cingulate cortex induces negative decision-making.
- Barker et al (2014). Individual Differences in Fear Potentiated Startle in Behaviorally Inhibited Children
- Borod et al (1997). Neuropsychological aspects of facial asymmetry during emotional expression: A review of the normal adult literature
- Blanchard et al (1997). Differentiation of anxiolytic and panicolytic drugs by effects on rat and mouse defense test batteries
- Blanchard & Blanchard (1990). An ethoexperimental analysis of defense, fear and anxiety.
- Bruno & Bertamini (2013). Self-Portraits: Smartphones Reveal a Side Bias in Non-Artists
- Calabrese (2010) L’arte dell’autoritratto: storia e teoria di un genere pittorico
- Carver & Harmon-Jones 2009. Anger is an approach-related affect: evidence and implications.
- Cisek (2021). Evolution of behavioural control from chordates to primates.
- Coan & Allen (2004). Frontal EEG asymmetry as a moderator and mediator of emotion.
- Corr (2008). The reinforcement sensitivity theory of personality.
- Cremers et al (2010). Neuroticism modulates amygdala-prefrontal connectivity in response to negative emotional facial expressions
- Crittenden et al (2015). Striosome–dendron bouquets highlight a unique striatonigral circuit targeting dopamine-containing neurons
- Davidson (1999). Neuropsychological perspectives on affective styles and their cognitive consequences
- Demaree et al (2005). Brain lateralization of emotional processing: historical roots and a future incorporating “dominance”
- d’Alfonso et al (2000). Laterality effects in selective attention to threat after repetitive transcranial magnetic stimulation at the prefrontal cortex in female subjects
- de Boer et al (2017). Untangling the neurobiology of coping styles in rodents: towards neural mechanisms underlying individual differences in disease susceptibility
- DeYoung et al (2007). Between Facets and Domains: 10 Aspects of the Big Five
- Depue & Collins (1999). Neurobiology of the structure of personality: Dopamine, facilitation of incentive motivation, and extraversion
- Elliot & Covington (2001). Approach and avoidance motivation
- Fetterman (2013). For Which Side the Bell Tolls: The Laterality of Approach-Avoidance Associative Networks
- Fitzgerald et al (2005). Error-related hyperactivity of the anterior cingulate cortex in obsessive-compulsive disorder
- Friedman et al (2015). A Corticostriatal Path Targeting Striosomes Controls Decision-Making under Conflict
- Goldstein et al (2009). Anterior cingulate cortex hypoactivations to an emotionally salient task in cocaine addiction
- Gray & McNaughton (1980). The Neuropsychology of Anxiety (first edition)
- Gray & McNaughton (2004). The Neuropsychology of Anxiety (second edition)
- Graeff (1994). Neuroanatomy and neurotransmitter regulation of defensive behaviors and related emotions in mammals.
- Gunturkun (2003). Adult persistence of head-turning asymmetry
- Helfinstein et al (2012). Approach-withdrawal and the role of the striatum in the temperament of behavioral inhibition
- Jaynes (1976). The Origin of Consciousness in the Breakdown of the Bicameral Mind
- Kagan et al (1984). Behavioral inhibition to the unfamiliar
- Kasai et al (2008). Evidence for acquired pregenual anterior cingulate gray matter loss from a twin study of combat-related posttraumatic stress disorder
- Koolhaas (2010). Neuroendocrinology of coping styles: towards understanding the biology of individual variation.
- Krack et al (2010) Deep brain stimulation: from neurology to psychiatry
- Le Doux (1994). Emotion, memory, and the brain
- MacNeilage (2007). Present status of the postural origins theory
- MacNeilage (1998). Towards a unified view of cerebral hemispheric specializations in vertebrates.
- Man et al (2017). Hierarchical brain systems support multiple representations of valence and mixed affect.
- McGilchrist (2009). The Master and His Emissary: The Divided Brain and the Making of the Western World
- McManus (2019). Half a century of handedness research: myths, truths; fictions, facts; backwards, but mostly forwards
- McNaughton & Corr (2004). A two-dimensional neuropsychology of defense: fear/anxiety and defensive distance.
- McNaughton et al (2016). Approach/Avoidance
- Miller (1944). Experimental studies of conflict.
- Mueller et al (2014). Dopamine modulates frontomedial failure processing of agentic introverts versus extraverts in incentive contexts
- Panksepp (1998). Affective neuroscience
- Perl et al (2019). Human non-olfactory cognition is phase locked with inhalation
- Pessoa et al (2019). Neural architecture of the vertebrate brain: implications for the interaction between emotion and cognition
- Pizzagalli (2011). Fronto cingulate dysfunction in depression: towards biomarkers of treatment response
- Robinson & Price (1982). Post-stroke depressive disorders: a follow-up study of 103 patients.
- Russel (1980). A circumplex model of affect
- Seo & Huh (2019). Automatic Emotion-Based music classification for supporting intelligent IoT applications.
- Shabel & Janak (2009). Substantial similarity in amygdala neuronal activity during conditioned appetitive and aversive emotional arousal
- Shadli et al (2021). Right frontal anxiolytic‑sensitive EEG ‘theta’ rhythm in the stop‑signal task is a theory‑based anxiety disorder biomarker
- Siegel & Victoroff (2009). Understanding human aggression: New insights from neuroscience
- Smillie (2008). What is Reinforcement Sensitivity? Neuroscience Paradigms for Approach-avoidance Process Theories of Personality
- Stuss & Alexander (2007). Is there a dysexecutive syndrome?
- Terzian (1964). Behavioral and EEG effects of intracarotid sodium amytal injection
- Wacker et al (2008). Is running away right? The behavioral activation-behavioral inhibition model of anterior asymmetry.
