Part Of: Demystifying Consciousness sequence
Content Summary: 800 words, 8 min read
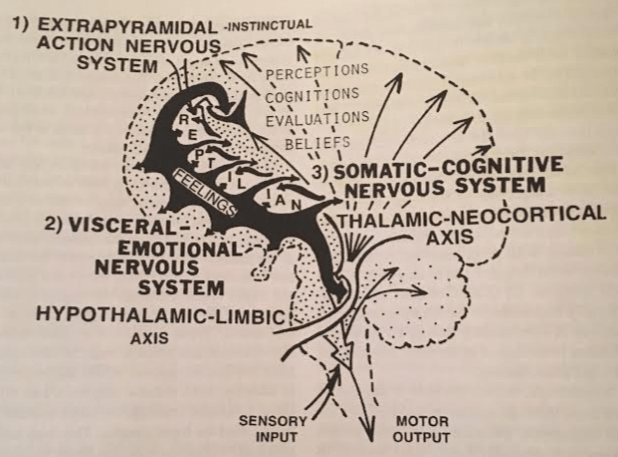
Existential Mode Generators
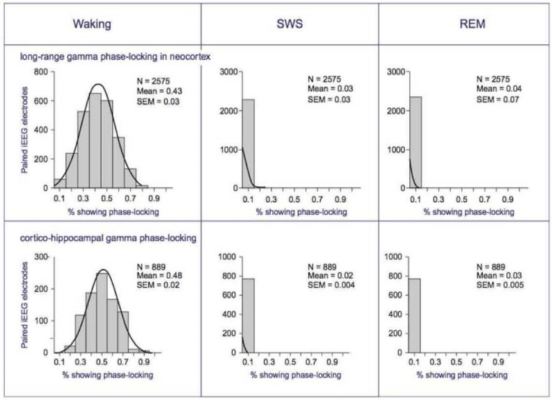
In Why We Sleep, we discussed sleep architecture diagrams. These diagrams show clear electrical differences between three existential modes: NREM (“sleeping”), REM (“dreaming”), and Consciousness.

While EEG excels at providing temporal resolution, it doesn’t provide much spatial information. Where does the brain construct these three modes?
To answer this, neuroscientists cut the brains of cats in half… literally. If you perform a Cerveau Isolé cut (slice above the midbrain), the top half’s electrical signature is NREM. If you do a Midpontine Pre-Trigeminal cut (slice below the midbrain), the top half’s electrical signature is NREM + Consciousness.

This evidence shows that existential modes are generated by different areas. Specifically:
- Sleep is induced by the diencephalon.
- Dreaming is initiated by the metencephalon.
- Consciousness is ignited by the mesencephon.
Neuroscientists now knew where to look! It was not long before they discovered the machinery that create consciousness, sleeping, and dreaming:

We now turn our gaze to the ascending reticular activating system (ARAS). “Reticular” is a word that means “web-like”, so the name roughly means “web-like ignition switch”. But before we do so, we need to turn our gaze to the relationship between cortico-thalamic (CT) radiations and consciousness.
Thalamus Anatomy & Function
We have also explained that the purpose of consciousness is to solve the binding problem: gluing together disparate adjectives into coherent nouns:
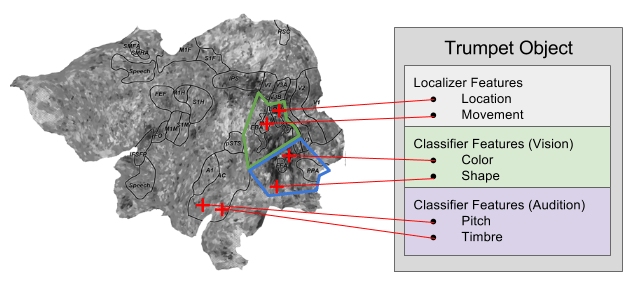
Consciousness creates the coherent objects of working memory by implementing phase binding, where object features are stitched together in distinct frequency bands, not unlike the radio in your car.
We have previously described the thalamus and cortex as dually innervating spheres, not dissimilar to a plasma globe:

And indeed, the nuclei within the thalamus tile the entire cortex:
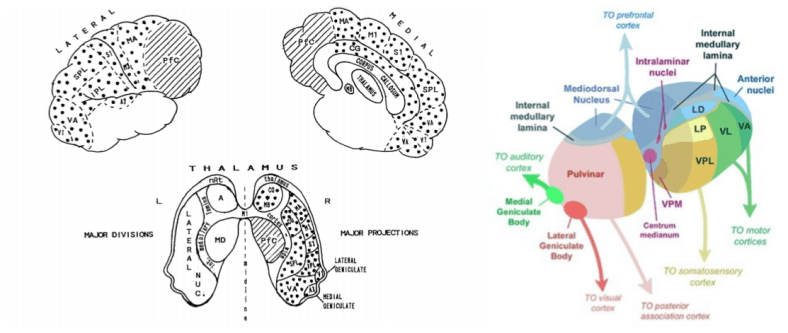
Note, however, that only some thalamic nuclei are specific (project to discrete patches of cortex). Nonspecific thalamic nuclei are also present, including the Intralaminar Nuclei (ILN) and Reticular Nucleus of the Thalamus (RNT).
These nonspecific nuclei are the principal components of the ERTAS system, and plausible candidates for the engine of consciousness.
Damage of specific nuclei produce loss of a particular modality. In contrast, lesions to nonspecific nuclei produces deep disturbances of consciousness. In fact, recent evidence suggests that such lesions perturb cortico-cortical information transmission.
The ERTAS Hypothesis
The ascending reticular activating system (ARAS) consists of a dense web of nuclei. Indeed, the word “reticular” means “web-like”. Parvizi, Damasio (2001) outline the more significant members of the system:

These nuclei project to the following three sites:
- Reticular Nucleus of the Thalamus (RNT), a sheet that sits on top of the thalamus.
- Intralaminar Nuclei (ILN), which are embedded deep within the thalamus.
- Basal Forebrain, which receives & distributes several neurochemical systems.
These structures in turn route information flowing to cortex:
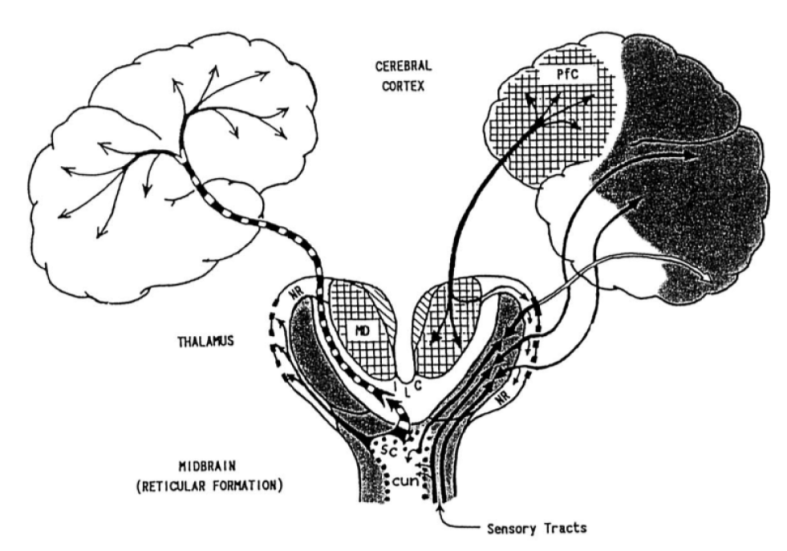
The extended reticular-thalamic activating system (ERTAS) hypothesis connects the ARAS system with the phase binding interpretation of the cortico-thalamo-cortical reentrant loop. One hypothesis, adapted from Newman (1999), has three theses:
- ILN performs phase binding (and thus, the consciousness generator).
- RNT implements selective attention.
- Basal Forebrain provides visceral “body-relevant” information.

More recent research has corroborated the role of the ILN in phase binding, and expanded its scope. Saalmann (2014) notes that the ILN seems to participate in a larger group of higher-order nuclei which each manage information within more constrained parts of cortex. The anterior ILN seems more related to oculomotor processes; the posterior deals with the multimodal integration of different sense data.
One unexpected recent finding has been that lesions of “higher-order nuclei” such as the ILN seem to perturb cortico-cortical information transmission. This underscores the need to understand interactions between the CTC Loop and other reentrant loops.

The Role of The Claustrum
The claustrum is a tiny sheet of gray matter suspended between thalamus and cortex. However, it receives information from essentially the entire cortex:

Given that the purpose of consciousness is to integrate cortical information, the anatomical position of the claustrum is suggestive.
Recent anatomical evidence has only strengthened the case for claustrum promoting consciousness:
- Koubeissi et al (2014) is a case study where they were electrical stimulation of the claustrum induced loss of consciousness (!).
- Chau et al (2015) announced evidence that correlate claustrum lesions with the duration, but not the frequency, of loss of consciousness.
- Wang et al (2016) conclusively proved that the claustrum has reciprocal connections everywhere in cortex.
- Reardon (2017) announced the discovery of a single neuron whose dendrites encircled the entire brain (image credit)
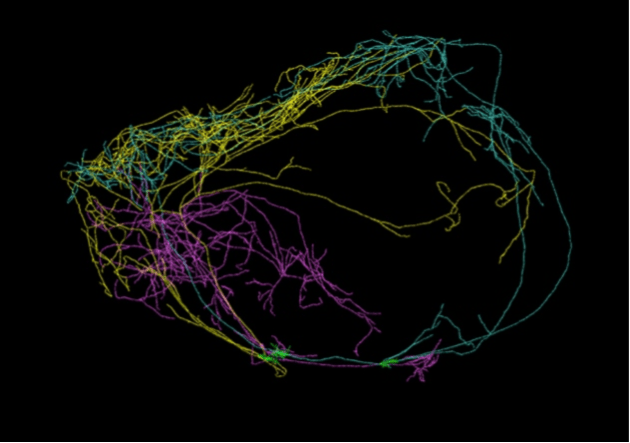
These data are suggestive. However, it will be some time before we know enough to integrate claustrum function within the ERTAS system.
Until next time.
Related Works
- Chau et al (2015). The effect of claustrum lesions on human consciousness and the recovery of function
- Crick, Koch (2005). What is the function of the claustrum?
- Koubeissi et al (2014). Electrical stimulation of a small brain area reversibly disrupts consciousness
- Newman (1999). Putting the puzzle together: towards a general theory of the neural correlates of consciousness
- Parvizi, Damasio (2001). Consciousness and the brainstem
- Reardon (2017). A giant neuron found wrapped around entire mouse brain.
-
Wang et al (2016). Organization of the connections between claustrum and cortex in the mouse