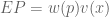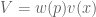Part Of: Demystifying Ethics sequence
Content Summary: 1500 words, 15 min
Motivations
Ethics discussions can occur at different levels of abstraction. The most concrete ethical discussions involve discussing specific topics; e.g., “is abortion moral”? These scenarios are called applied ethics.
Spend enough time wrestling with applied ethics, and you’ll start wondering what all of these moral intuitions have in common. An ethical theory is an answer to the question “what makes a thing good or bad”?
Meta-ethics answers a yet broader set of questions. What is right and wrong, anyways? Are moral beliefs objectively true in the same sense that the Earth orbits the sun? Or is my condemning an unethical act merely expressing an aesthetic preference (“yuck!”)?

Image Credit
Today we conduct a whirlwind tour of ethical theories, touching on the most influential theories. Ethical theories are normative: they speak to what moral conclusions ought to be. However, we will see later how these theories also sometimes imperfectly anticipate the emerging science of moral cognition.
We will discuss the following:
- Divine Command Theory
- Natural Law Theory
- Virtue Theory
- Deontology
- Utilitarianism
The first three theories are the oldest, and have close ties to religious doctrine. The last two are the most well-known theories these days.
Divine Command Theory
Historically, religions have had a good deal to say about right and wrong. One of the oldest ethical theories is Divine Command Theory, which states:
Moral behaviors are those that are commanded (or willed) by the divine.
This theory was originally proposed in the context of Greek mythology. But since it doesn’t say much about the specific character of the divine, it was adopted by Augustine and some (but not all) modern theologians.
One of the strongest criticisms to Divine Command Theory is the Euthyphro Dilemma, which goes as follows:
- Is an action ethical because God commands it?
- Or does God command it because it is ethical?
Clearly, we must affirm one of these options. But which one? Either “horn” of the dilemma extracts painful concessions:
- On the first option, moral content becomes arbitrary. If God changes his mind about moral facts, our moral universe is turned upside down.
- On the second, God becomes a middleman who conveys independent ethical facts. Why can’t we simply learn moral truths direct from the source?
These days, most philosophers find Divine Command Theory problematic. But there are other ethical theories available to the faithful.
Natural Law
Another approach to ethical theories, championed by Thomas Aquinas, is Natural Law Theory. Moral truths are grounded as follows:
Moral behaviors are those which promote, and do not detract from, a basic good.
A basic good is injected into human nature by God. Here are three such lists, generated by a Catholic, a Muslim, and a philosopher:
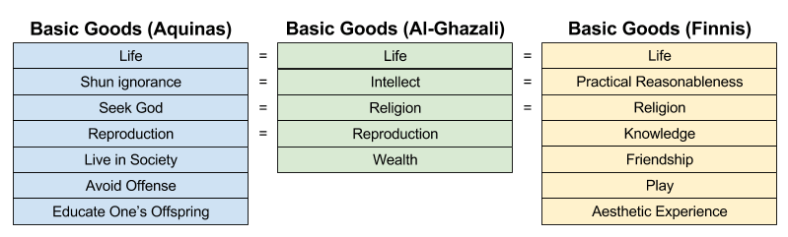
Ethical actions promote the basic goods; immoral actions detract from the basic goods. This, incidentally, is why Catholics oppose all forms of birth control: they detract from the basic good of Reproduction.
How did these philosophers generate their lists of basic goods? By observing nature. For example, Life and Reproduction usually make the cut because humans naturally prioritize survival and reproductive success (this is a consequence of natural selection).
However, there is a conceptual difficulty in deriving basic goods from nature. This was clearly expressed by Hume’s Law, also called the is-ought gap: what logical procedure could possibly be used to infer normative statements (an “ought”) from descriptive facts of the words (an “is”)?
Virtue Theories
Unlike its competitors, virtue theory doesn’t spend much time telling you what to do (moral behaviors). Instead, people are the subject of moral truths. Behaviors are only good or evil insofar as they enhance or corrupt a person’s character.
Aristotle presented these ideas in Nichomachean Ethics. There he describes ethical behavior as a way to achieve eudaimonia (human flourishing or “the good life”).
A life of eudaimonia is a life of striving. It’s a life of pushing yourself to your limits, and finding success. A eudaimonistic life will be full of the happiness that comes from achieving something really difficult, rather than just having it handed to you.
Aristotle also believed that moral knowledge is learned not by navel-gazing, but instead comes from social interactions. Specifically, good character is developed by spending time with virtuous people. This theory is where the idea of a role model comes from.
So let’s get down to business. In virtue theory, moral facts are grounded as follows:
Moral people are those who have character; that is, they possess basic virtues.
Aristotle identified twelve basic virtues: Courage, Moderation, Flexibility, Cleanliness, Generosity, Ambition, Patience, Truthfulness, Wittiness, Friendliness, Modesty, and Righteous Indignation. A good person nurtures these virtues.
The Golden Mean provides some insight into how these virtues can be refined over time. A person can respond too strongly with a virtue (excess), or too weakly (deficiency). For example, one must not have too much courage, nor too little:

Virtue ethics is surprisingly robust to philosophical criticism. However, one scientific concern has been gaining traction recently. Empirical research in the field of situational psychology increasingly suggests that there is no such thing as character traits.
Make no mistake, seeing vices and virtues in others is a universal human instinct. But there is no guarantee that mental software works correctly (e.g., see how the brain lies to itself about visual experience). And the fact of the matter is, character information contributes zero predictive power over how a person will respond to a given situation.
Duty Theories
The first ethical theories that isn’t explicitly theistic, Immanuel Kant’s deontological (“knowledge of what is proper”) system purports to result from pure reason.
Moral behaviors are those which obey the categorical imperative.
For Kant, the categorical imperative is a procedure by which a person can discover the rigid contents of the moral universe. He presents four perspectives that independently arrive at these moral truths; the two most influential of which are:
- The universalizability principle instructs us to act only if you also desire that your action should become a universal law. Lying is wrong because if lying becomes universalized, meaningful communication becomes impossible.
- The principle of humanity instructs us to act in such a way that you treat humanity (both yourself or someone else), always also as an end and never simply as a means. This principle underlies the modern idea of human rights, and is leveraged by Nozick’s brand of libertarianism.
The categorical imperative promotes a kind of moral absolutism: if actions are right or wrong intrinsically, their moral status is invariant to the consequences of an action. And this inflexibility can produce counterintuitive judgments.
For example, consider a house shielding a Jewish family from the Nazis. If a soldier comes to the door asking about its residents, most of use think it would be morally acceptable to lie. Kant, however, claims that lying is wrong in all situations, even ones such as these.
Consequentialist Theories
Deontological theories are invariant to behavioral outcome. But there are many moral situations where outcome seems to matter. Consider the trolley problem, where a trolley is currently barreling down a track and should kill five people unless diverted. However, on the other track one person has been similarly demobilized:
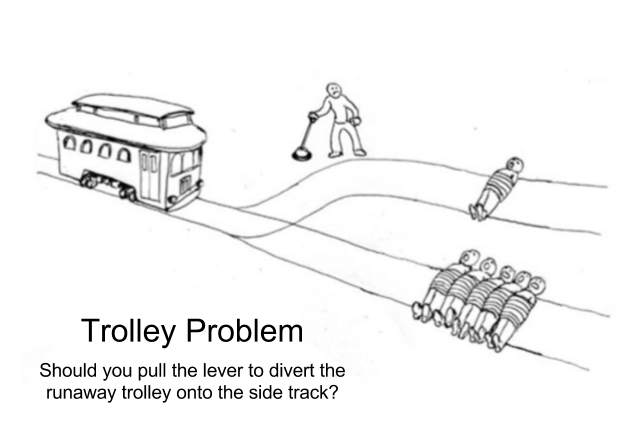
Kant would say that pulling the lever is simply murder. That five people die is regrettable, but their death does not incriminate the person at the switch. If the person does divert the trolley, the guilt of the single person’s death is on his head.
However, most people think that the number of deaths – the consequence of an action – matters. Consequentialist theories make this intuition explicit:
Moral behaviors are those that bring the most good to the most people.
Consequentialist theories vary on how they define “the most good” (personal well-being) and “the most people” (societal well-being).
For example, classical utilitarianism, advocated by Jeremy Bentham and John Stuart Mill, which takes the following stances:
- Personal well-being originates from hedonism: actions are moral if they increase pleasure and/or decrease pain.
- Societal well-being is taken as the sum total of the well-being of constituent individuals.
Many people condemn utilitarianism for its hedonism (“don’t immoral behaviors often feel pleasurable?”). However, this criticism loses some of its force when you remember to distinguish higher pleasures (eg., aesthetic experience) versus baser pleasures.
The problem of figuring out societal well-being, how to optimize outcomes for communities, is much more challenging. The repugnant conclusion, aka the mere addition paradox, is a particularly damning criticism. A large society with barely tolerable quality of life (e.g., urban sweat shops) doesn’t feel morally equivalent to happier, smaller societies.
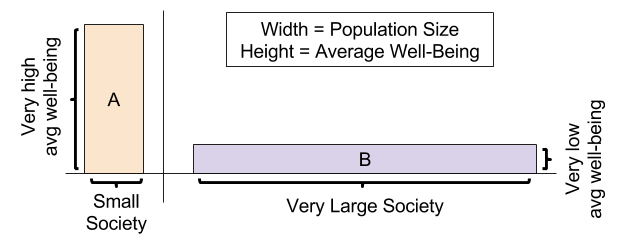
Thus, maximizing total well-being may be problematic. But different problems emerge if we instead optimize against average well-being. To this day, designing aggregation functions to compute societal well-being remains an essentially unsolved problem.
Takeaways
Today, we discussed the following theories:

There exist other theories besides these, of course. For example, Hobbes’ contractualism makes important contributions to the conversation.
To date, no ethical theory has emerged as a clear favorite. A recent survey of professional philosophers reveals considerable support for three of the above theories:
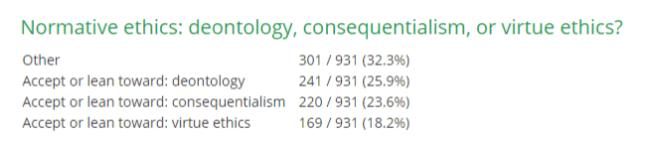
Next time, we will explore whether we should view these normative theories as competitors, or complementary solutions to different concerns. Until then.















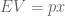



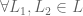 either
either  or
or  or
or 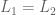
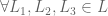 if
if  then
then 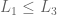
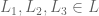 then
then  s.t.
s.t. 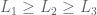 iff
iff 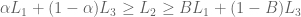
 over
over 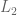 , mixing both with the same third lottery
, mixing both with the same third lottery  in the same proportion α must not reverse that preference—adding identical “padding” is irrelevant to the choice.
in the same proportion α must not reverse that preference—adding identical “padding” is irrelevant to the choice.


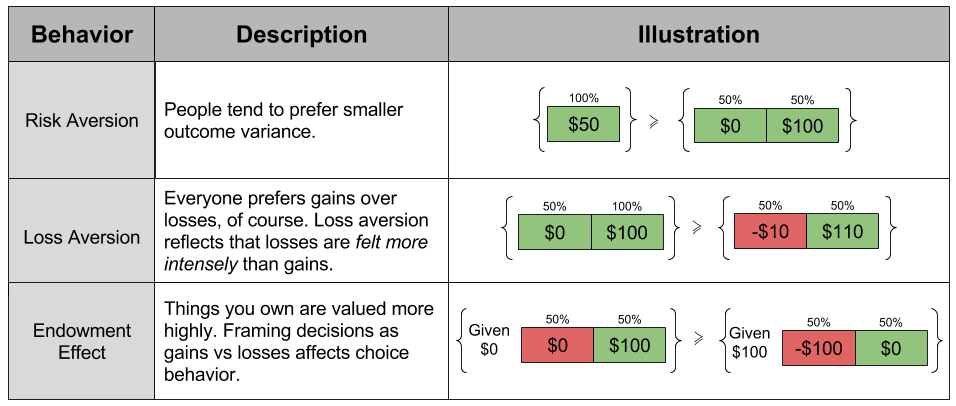
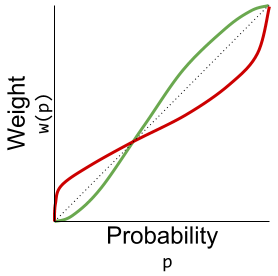
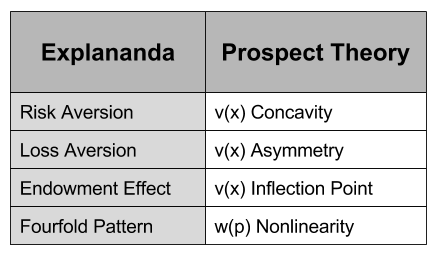
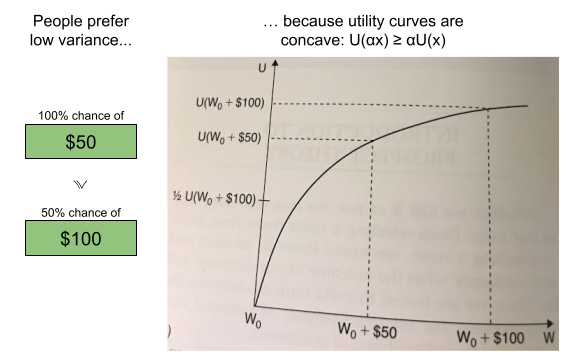
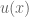 concavity to explain risk aversion? Prospect theory takes this approach yet further, and seeks to explain all of the above behaviors using a more complex shape
concavity to explain risk aversion? Prospect theory takes this approach yet further, and seeks to explain all of the above behaviors using a more complex shape represent our updated notion of utility. We can define expected prospect
represent our updated notion of utility. We can define expected prospect  of a function as probability multiplied by the value function
of a function as probability multiplied by the value function

 :
:
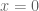 , captures the status quo. Thus, the reference point allows us to differentiate gains and losses, thereby producing the endowment effect.
, captures the status quo. Thus, the reference point allows us to differentiate gains and losses, thereby producing the endowment effect.
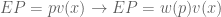
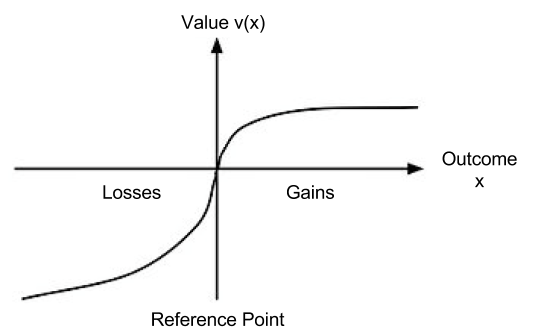
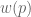 has the following shape:
has the following shape: